Soil mineral N dynamics beneath mixtures of leaves from legume and fruit trees in Central Amazonian multi-strata agroforests
Dinâmica do nitrogênio mineral no solo em misturas de folhas de leguminosas arbóreas e de fruteiras em sistemas agroflorestais multiestratificados na Amazônia Central
Carol Melanie Schwendener1, Johannes Lehmann1*, Marco Rondon2, Elisa Wandelli3, and Erick Fernandes4
1 909 Bradfield Hall, Department of Crop and Soil Sciences, Cornell University, Ithaca, NY 14853, USA. e-mail: cms67@cornell.edu *corresponding author: cl273@cornell.edu, (1) 607 254-1236
2 International Center for Tropical Agriculture (CIAT), A. A. 6713, Cali, Colombia, m.rondon@cgiar.org, (57) 2 445-0000
3 Empresa Brasileira de Pesquisa Agropecuaria (Embrapa) – Amazonia Ocidental, C. P. 319 69011-970 Manaus-AM, Brazil, elisa@cpaa.embrapa.br, (55) 92 621-0412
4 Agriculture and Rural Development, The World Bank, 1818 H Street, Washington, D.C. 20433, efernandes@worldbank.org, (1) 202 473-1292
ABSTRACT
Long term applications of leguminous green mulch could increase mineralizable nitrogen (N) beneath cupuaçu trees produced on the infertile acidic Ultisols and Oxisols of the Amazon Basin. However, low quality standing cupuaçu litter could interfere with green mulch N release and soil N mineralization. This study compared mineral N, total N, and microbial biomass N beneath cupuaçu trees grown in two different agroforestry systems, north of Manaus, Brazil, following seven years of different green mulch application rates. To test for net interactions between green mulch and cupuaçu litter, dried gliricidia and inga leaves were mixed with senescent cupuaçu leaves, surface applied to an Oxisol soil, and incubated in a greenhouse for 162 days. Leaf decomposition, N release and soil N mineralization were periodically measured in the mixed species litter treatments and compared to single species applications. The effect of legume biomass and cupuaçu litter on soil mineral N was additive implying that recommendations for green mulch applications to cupuaçu trees can be based on N dynamics of individual green mulch species. Results demonstrated that residue quality, not quantity, was the dominant factor affecting the rate of N release from leaves and soil N mineralization in a controlled environment. In the field, complex N cycling and other factors, including soil fauna, roots, and microclimatic effects, had a stronger influence on available soil N than residue quality.
KEYWORDS: Theobroma grandiflorum, Gliricidia sepium, Agroforestry, Mineral N, Green mulch
RESUMO
Aplicações a longo prazo de leguminosas como adubo verde podem aumentar o nitrogênio (N) mineralizável sob árvores de cupuaçu em solos pouco férteis e ácidos (Ultisols e Oxisols) da Bacia Amazônica. Entretanto, a baixa qualidade da liteira de cupuaçu pode influênciara liberação de N do adubo verde e a mineralização deste no solo. Neste estudo foram comparados o N mineral, N total, e o N da biomassa microbiana sob árvores de cupuaçu cultivadas em dois sistemas agroflorestais, ao norte de Manaus, Brasil, as quais receberam diferentes aplicações de adubo verde sob sua liteira natural durante sete anos. Para testar as interações entre o adubo verde e a liteira de cupuaçu, folhas secas de gliricídia e ingá foram misturadas com as folhas senescentes de cupuaçu, distribuídas na superfície de um solo Oxisol, e incubadas em casa de vegetação durante 162 dias. A decomposição das folhas, a liberação de N e mineralização do N no solo foram periodicamente mensurados nos tratamentos de mistura de liteira de diferentes espécies e comparados com as aplicações de liteira de apenas uma espécie. O efeito da biomassa de leguminosas e da liteira de cupuaçu no N mineral do solo foi aditivo, indicando que o uso de adubação verde em plantas de cupuaçu pode ser baseado na dinâmica do N em cada espécie usada como adubo verde. Os resultados demonstraram que a qualidade do resíduo, e não a quantidade, foi o principal fator que influenciou a taxa de liberação de N das folhas e a mineralização deste no solo, em ambiente controlado. No campo, o complexo ciclo do N e outros fatores tais como a fauna do solo, raízes e os efeitos do microclima, tiveram uma influência mais forte na disponibilidade de N no solo do que a qualidade d o resíduo vegetal.
PALAVRAS-CHAVE: Theobroma grandiflorum, Gliricidia sepium, Sistemas agroflorestais, N mineral, Adubação verde
INTRODUCTION
Prunings from leguminous trees provide an accessable organic nitrogen (N) source for small-scale agroforestry farmers on the acid infertile Oxisol and Ultisol soils that dominate the Amazon Basin (Sanchez et al., 1982). In alley cropping and shade agroforestry systems N applied with prunings can amount to 136-240 kg N ha-1 y-1 on infertile soils (Kang et al., 1981; Russo & Budowski, 1986). 37-92% of the N in gliricidia (Gliricidia sepium) prunings can be fixed from atmospheric N2 (Giller, 2001). Although applications of prunings from leguminous species can increase crop yields ( Kang et al., 1981; Haggar et al., 1993; Mulongoy et al., 1993) and organic soil N and N mineralization (Isaac et al., 2003), often only 10-20% or less of the N added in the prunings is recovered by the first subsequent annual crops (Haggar et al., 1993; Palm, 1995). Much of the N not used by crops remains in undecomposed material or moves to the readily mineralizable fraction of soil organic matter (Haggar et al., 1993), while significant amounts can be leached to the subsoil (Schroth et al., 1999), or lost through volatilization (Palm, 1995). Agroforestry tree species have the potential to benefit from residual N or priming effects of long term green mulch applications (Cadisch et al., 1998) due to their longer growing periods, more established root systems and a lower need for nutrient synchrony than annual crops.
Extensive research has shown that applications of legume tree prunings increase available N in agroforestry systems, but no studies have focused specifically on application in tree cropping systems. Cupuaçu (Theobroma grandiflorum (Wild. ex Spring) Schumann) is a common fruit tree species in agroforestry systems in the Amazon Basin that produces a low quality litter (high C:N ratio and high lignin contents) (McGrath et al., 2000; Uguen, 2001). Applications of high quality prunings, such as gliricidia, could increase the nutrient release from cupuaçu litter. It has been shown that cupuaçu litter can immobilize P from higher quality litter and decrease available P in the soil (McGrath et al., 2000), but little is known about whether low quality cupuaçu litter decreases soil N mineralization, green mulch decomposition and N release.
Green mulch and litter decomposition and N mineralization rates have been related to various intrinsic chemical properties of residues and ratios of the parameters, including initial N, lignin, and polyphenol contents, C/N ratio (Constantinides and Fownes, 1994; Handayanto et al., 1997; Lehmann et al., 1995; Mafongoya et al., 1998a; Palm & Sanchez, 1991; Tian et al., 1992). These predictors of residue decomposition and N mineralization rates can be altered by mixing different quality leaves to regulate N release (Handayanto et al., 1997). Although much work has been done on the residue quality parameters to predict N release and decomposition, only a few studies have looked at interactions of multi-species mulch or litter mixtures (Handayanto et al., 1997; Mafongoya et al., 1998b; McGrath et al., 2000) and none specifically at low-quality litter from tree crops such as cupuaçu mixed with high-quality legume prunings such as gliricidia and inga (Inga edulis).
The objectives of this research were 1) to determine whether green mulch quality and quantity increase soil mineral N beneath cupuaçu in a controlled environment and in an agroforestry system; 2) to assess whether cupuaçu litter decreases soil N mineralization and N release by green mulch.
MATERIAL AND METHODS
SITE DESCRIPTION
The project site is located in central Amazonia at the Empresa Brasileira de Pesquisa Agropecuaria – Centro da Pesquisa Agroflorestal (EMBRAPA-CPAA) research station 53 km north of Manaus, Brazil along the highway BR-174. The soil is a fine, isohyperthermic, Xanthic Hapludox (U.S. Soil Survey Staff, 1998) or Ferrasol (FAO-UNESCO, 1990). The annual average rainfall is 2500 mm per year with a 2-3 month dry season, and the mean annual temp is 26 °C. Between 1991 and 1992, three 50 m x 60 m plots each of a palm-based agroforestry system (Agrosilvicultural I - ASI) and a fruit-tree based home garden system (Agrosilvicultural II - ASII) were established on abandoned, degraded pastureland. Details on site preparation and establishment can be found in Fernandes et al. (submitted). Species composition of each system is shown in Figure 1.
Both systems were bordered with around 90 gliricidia trees spaced two meters apart and intercropped with cupuaçu trees. In ASII, the cupuaçu trees were planted in two 13-tree rows in six by two meter spacing, for a total of 26 cupuaçu trees with 25 intercropped inga trees, also spaced six by two meters apart. Agrosilvicultural system I had 99 cupuaçu trees spaced six by two meters apart and intercropped with açaí (Euterpe oleracea) trees only or with açai and capoeirão (Colubrina glandulosa) trees (Figure 1). Gliricidia prunings were applied two to three times annually to cupuaçu trees in both systems for seven years. In ASII, inga prunings were applied to cupuaçu trees in addition to the gliricidia prunings two to three times annually for seven years. The lower density of cupuaçu trees in ASII and similar total gliricidia pruning biomass resulted in individual trees in ASII receiving higher rates of gliricidia prunings than trees in ASI. The addition of inga prunings to trees in ASII further increased the difference in green mulch applications between the two systems. The nitrogen input per hectare was 1.46 times greater in ASII than in ASI (16.8 and 24.5 kg N ha-1 y-1 for ASI and ASII, respectively) (EMBRAPA-CPAA, 1997), or 5.6 times greater per cupuaçu tree (approximately 0.17 and 0.94 kg tree-1 for ASI and ASII, respectively). At the time of sampling, the cupuaçu trees were 10 years of age and green mulch applications had been occurring for seven years.
SAMPLING
Fresh gliricidia prunings were applied to four randomly selected cupuaçu trees (excluding edge trees) within each block in ASI and ASII. Prunings were applied at a rate of 10 kg fresh weight per tree (3 kg leaves, 7 kg branches), branches placed on top to hold the leaves in place, within a 0.5 m radius of the tree trunks. The application rate and method was similar to past applications in the agroforestry systems. One week after application, during which at least one rainfall occurred, the top 0-10 cm of soil was sampled at 0.2 m and 1 m from the base of the tree. The 0.5m distance represented the area that always received inga and/or gliricidia prunings, while the 1 m location was primarily covered with cupuaçu litter and some litter from adjacent species. The soil was sampled at 2 locations for each distance from each tree and composited for each distance for each treatment and replicate, resulting in a total of 12 samples (two treatments, two distances, three replicates). Plots were sampled in July 2001 at the end of the rainy season and again in November 2001 at the end of the dry season.
GREENHOUSE INCUBATION
The substrate soil was the same Xanthic Hapludox (Ferrasol) on which the agroforestry systems were established, but it was collected beneath an 8-year old secondary forest adjacent to the ASI and ASII plots. Soil was collected from 7.5 cm to 15 cm depth and the large roots and charcoal pieces were excluded to minimize initial organic matter content and facilitate detection of treatment effects. Aggregates greater than 2 cm were broken apart to increase homogeneity of the soil without complete destruction of macro-aggregation in the soil. The soil had the following characteristics: pH (in water) 4.2; 1.6 g kg-1 N (Kjeldahl), 21.2 g kg-1 C (Walkley-Black), 1.8 g kg-1 available P (Mehlich-1), 18.3 g kg-1 available K (1M KCl), 22.7 g kg-1 Ca (1M KCl), and 11.2 g kg-1 Mg (1M KCl).
Senescent cupuaçu leaves, collected during the four weeks prior to application, and fresh gliricidia, and inga leaflets were cut into 1 cm2 pieces, the internodes of gliricidia and inga to 2 cm, dried to a constant weight at 55-65 ºC, and stored in dry conditions. The application rates included internodes and leaves at the same ratio found in leaves of the plants. Initial C and N contents of the leaves for gliricidia were 33.6 mg g-1 N, 421 mg g-1 C, for inga 28.4 mg g-1 N and 436 mg g-1 C and for cupuaçu 10.1 mg g-1 N and 361 mg g-1 C.
Exactly 150 g of fresh soil were weighed into 500 mL plastic cups, resulting in a surface area of 50 cm3. Care was taken to prevent drying of the soil. The samples were placed in a randomized block design in a greenhouse protected from rain and direct sun. The soils were left undisturbed for three weeks prior to application of litter treatments to allow microbial activity to stabilize. Daytime soil temperatures averaged 32-36 ºC. Soil moisture was maintained at 66% field capacity, except at 12 days prior to each collection when a rainfall was simulated and the moisture content was raised to 80% field capacity. This flush of water was intended to facilitate movement of N through the litter layer to the soil.
Treatments consisted of single and mixed species litter applications and a bare soil control. Gliricidia, inga, and cupuaçu were applied separately, as single species. Multiple species treatments were composed of the following combinations: gliricidia + inga, cupuaçu + gliricidia, cupuaçu + gliricidia (high rate), cupuaçu + gliricidia + inga and cupuaçu + gliricidia + inga (high rate). Gliricidia and inga leaves were applied at a low rate in all treatments, except those indicated as high rate. The high and low application rates of gliricidia and inga, 2 Mg ha-1 (1.0 g cup-1), 10 Mg ha-1 (5.0 g cup-1) and 2.6 Mg ha-1 (1.3 g cup-1), 13 Mg ha-1 (6.5 g cup-1), respectively, were selected to extend the range of application rates in the field. The application rate for cupuaçu litter of 4 Mg ha-1 (2.0 g cup-1) was based on standing cupuaçu litter and litter production in agroforestry systems similar to ASI (Uguen, 2001). The total N applied per cup as litter differed for each treatment as follows: 20.2 mg (cupuaçu), 33.6 mg (gliricidia), 36.9 mg (inga), 70.5 mg (gliricidia + inga), 53.8 mg (cupuaçu + gliricidia, low rate), 188.2 mg (cupuaçu + gliricidia, high rate), 90.8 mg (cupuaçu + gliricidia + inga, low rate), and 372.8 mg (cupuaçu + gliricidia + inga, high rate). All leaf combinations were mixed and surface applied to the soil with an amount of water equivalent to the water weight lost during drying. There were five replicates of each treatment and the control. Soil and litter were destructively sampled at 7, 29, 50, 98, and 162 days after litter application.
LABORATORY ANALYSES
Soils from the field and greenhouse incubation were analyzed for mineral N, total N and C, and field soil samples were also analyzed for microbial biomass C and N. Mineral N was extracted from 25 g fresh soil within two hours after collection with 75 mL of 1N KCl for 0.5 hour on a horizontal shaker at the highest speed. After decanting, the supernatant was pipetted and stored below 0 °C until it was analyzed for ammonium and nitrate on a Skalar continuous flow analyzer. Results are reported per dry weight soil, determined by drying at 105 ºC for 24 hours. Net interactive effects on decomposition, litter N release, and soil mineral N were estimated by summing values for single species treatments and statistically comparing them to the equivalent low rate mixed species treatment values. The control soil mineral N value was subtracted from all treatments to discount the mineralized N in the soil that was not a direct result of litter application. Microbial biomass N was measured by chloroform fumigation-extraction (Brookes et al., 1985) adapted for acid soils. The remaining soil was dried and ground to 2 mm prior to N and C analysis. Remaining plant material was cleaned by gentle brushing, dried for 48 hours at 65-70 °C, weighed, ground, and analyzed as a mixture for N and C. Plant and soil N were analyzed by Kjeldahl digestion, and carbon was analyzed by the Walkley-Black method (Silva, 1999).
STATISTICAL ANALYSES
Statistical analysis of the data was performed with regressions and 1-way and 2-way ANOVAs in Minitab 13 (Minitab, 2000). Where p < 0.05, means were compared using Fisher’s least significant difference test.
RESULTS
GREENHOUSE INCUBATION - LITTER DECOMPOSITION
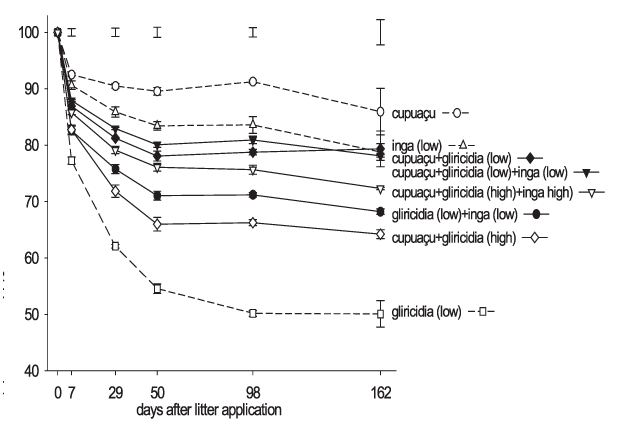
The gliricidia leaves applied in isolation decomposed more rapidly and completely of all single and mixed species litters as a percent of the dry weight applied. The cupuaçu litter applied alone decomposed the slowest during the 162-day experimental period (Figura 2). Among the multiple species leaf mixtures, the cupuaçu + gliricidia leaves applied at a high rate decomposed significantly faster and to a greater extent than other mixed species litters. The leaf mixtures with high rates of green mulch decomposed significantly faster than equivalent mixtures with low green mulch application rates; however, only the high and low rate cupuaçu + gliricidia leaf mixtures differed significantly in total decomposition with the high rate decomposing to a greater extent during the experimental period. Total leaf decomposition of species applied together was not significantly different from the total decomposition of the species when applied singly.
GREENHOUSE INCUBATION - N RELEASE FROM LITTER
The gliricidia leaves released the highest percent N of all single and mixed species litters, while the cupuaçu and inga leaves released the least percent N (Figura 3a). The percent N release by inga was only significantly greater than the percent N released by cupuaçu at 50 days after application. The cupuaçu + gliricidia (high rate) leaf mixture initially immobilized less than 1% of the nitrogen before quickly releasing more than 60% and then immobilizing about 10% N through to 162 days after application (Figura 3b). This release pattern was different from that of the other cupuaçu and green mulch litter mixtures, which had an initial rapid N release leveling at or prior to 162 days. Both treatments with cupuaçu + gliricidia + inga leaves released N more slowly than the treatments with only cupuaçu + gliricidia leaves. Despite different release patterns, there was no significant difference among the mixtures in total N released 162 days after litter application.
GREENHOUSE INCUBATION - SOIL N
Net mineral N accumulation in the soil was better correlated with the amount of gliricidia N applied (r = 0.498; n = 219; p <0.001) than total plant N applied (r = 0.204; n = 219; p <0.001). Treatments with the higher gliricidia application rate had significantly greater soil mineral N levels than the treatments with less gliricidia applied (Fig. 4). Cumulative N mineralized as a percent of the N applied in the litter mixture was lower in the cupuaçu +gliricidia + inga treatments compared to the cupuaçu +gliricidia treatments when high and low application rates were averaged (Figura 4, inset). Calculations to determine net interactions showed no significant differences in soil mineral N when the leaves were applied separately or in combination. The presence of cupuaçu leaves with inga and/or gliricidia leaves did not reduce soil mineral N compared to identical treatments without cupuaçu. The mineral N in the gliricidia + inga treatment was not significantly different from the treatment with only gliricidia leaves. There was no treatment effect on total soil N during the 162-day experimental period.
Mineral N, total N, and microbial biomass C and N did not differ between ASI and ASII at either distance from cupuaçu trees, however, nitrate was higher in ASII, due to higher nitrate levels 0.20 m from the base of the tree. In both systems, soil mineral N was greater at 0.2 m from the tree than at 1 m, but distance had no effect on total N or microbial biomass C or N.
DISCUSSION
As expected, residue quality, defined by the C:N ratio, was the dominant factor affecting the rate of N release from leaves and soil N mineralization in the greenhouse incubations. This was shown by the stronger positive correlation between gliricidia N applied and soil mineral N than between total mulch N applied and soil mineral N. Further evidence was the more rapid, but not greater, N release from litter mixtures with gliricidia leaves. Nitrogen mineralization in the soil as a percent of the N applied in the mulch decreased significantly when lower quality litter (inga) was included in the green mulch added to cupuaçu. Also, inga and cupuaçu leaves applied separately did not increase soil mineral N above bare soil values, and leaf mixtures with inga did not increase soil mineral N above that of similar mixtures without inga. This suggests that during the 162-day experimental period, inga and cupuaçu did not increase or decrease soil mineral N as is expected for litter species with decomposition half-lives greater than the study period (over 1 year for cupuaçu (McGrath et al., 2000) and 39 weeks for air-dried inga (Palm and Sanchez, 1990).
Soil mineral N levels were additive in the lower rate mixtures, and, contrary to expectations, no net interactions between the cupuaçu, gliricidia, and inga litters occurred. The presence of cupuaçu did not decrease net N mineralization or N release in the lower rate green mulch applications. The additive effect of legume biomass and cupuaçu litter on soil mineral N levels implies that recommendations for green mulch applications to cupuaçu trees can be based on N dynamics of individual green mulch species and crop N requirements. Soil mineral N quantities, however, cannot be extrapolated from lower application rates because the increase in soil mineral N was not proportional to the increase in applied N. This may be due to a larger litter layer with higher application rates, which decreased soil contact with the leaves and consequently slower decomposition (Henriksen and Breland, 2002; Wilson et al., 1986). The absence of net interactions does not preclude gross interactions between cupuaçu litter N and green mulch N, which were not studied here. Schwendener et al. (unpublished data) also found no net interactions between cupuaçu and gliricidia leaves, but with the use of 15N were able to confirm that gross interactions, including temporary immobilization by cupuaçu of N released by gliricidia, were occurring. An understanding of gross interactions among cupuaçu litter, green mulch and soil could facilitate the management of litter and green mulch N dynamics farmer field conditions.
In a homogenous soil and controlled environment, higher quantities of high quality litter (i.e. gliricidia) increased soil mineral N; under field conditions in the agroforests, however, other factors had a greater effect on soil mineral N than the quality of the green mulch. Soil mineral N and total soil N were expected to be higher in ASII due to accumulation of readily mineralizable N fraction in soil organic matter during the seven years of higher green mulch applications, but soil mineral N, microbial biomass N, and total N did not differ between the two agroforestry systems. Temporal variability of mineral N and the short sampling period of this study made it difficult to examine N mineralization rates which may have better reflected the greater accumulation of a mineralizable N fraction as a result of long term green mulch applications (Haggar et al., 1993). Other factors present in the field and absent in a controlled experiment, such as soil fauna (Hofer et al., 2001; Tian et al., 1992; Vohland and Schroth, 1999), roots (Lehmann et al., 2001; Schroth et al., 1999) and microclimatic effects such as increased shade and moisture (Giller, 2001; Rao et al., 1997), may have had larger effects on soil mineral N than the application of green mulch and may have affected soil mineral N differently in ASI than in ASII. For instance, macrofauna may have been more abundant in ASI due to the close proximity of cupuaçu to peach palm and consequently increased decomposition of the higher quality litter (Vohland and Schroth, 1999). Alternatively, the additional gliricidia N added to ASII could have been removed from the system through increased export of fruit (which was higher in ASII (Wandelli, pers. com.)) or lost via leaching of nitrate to the subsoil (Schroth et al., 1999). Higher mineral N closer to the trunk in both systems suggests that in addition to higher rates of gliricidia prunings at that location, increased shade and thicker litter layer contribute to a favorable microclimate and increased N mineralization (Babbar and Zak, 1994).
The strong influence of gliricidia on soil N mineralization in the greenhouse study demonstrates the importance of the quality of N applied, rather the quantity of N applied as mulch for short term soil N availability. When referring only to higher quality mulch, quantity is also significant for increasing N mineralization. The lower quality inga mulch and cupuaçu litter did not interfere with net decomposition of and N release of gliricidia mulch and soil mineral N accumulation in the soil mulched with gliricidia. Although the low-quality and slow-decomposing inga and cupuaçu leaves did not contribute to soil mineral N in the short term, they could benefit soil fertility in ways not studied in this paper, by providing a litter layer for soil fauna, reducing erosion, increasing soil moisture availability, and controlling weeds. Laboratory studies on soil N availability are limited to green mulch management recommendations with respect to soil N availability. Final recommendations for green mulch applications should be based on field trials that include the N sources and sinks which contribute to the complex N cycling in agroforestry systems.
ACKNOWLEDGMENTS
This research was funded by NASA Large Scale Biosphere Atmosphere-Ecology (LBA-Eco) project (ND-04). The authors thank EMBRAPA and INPA Soil and Plant Laboratories for their assistance with soil and plant analyses.
LITERATURE CITED
Babbar, L.I.; Zak, D.R. 1994. Nitrogen cycling in coffee agroecosystems: net N mineralization and nitrification in the presence and absence of shade trees. Agriculture, Ecosystems, and Environment, 48(2): 107-113.
Brookes, P.C.; Landman, A.; Pruden, G.; Jenkinson, D.S. 1985. Chloroform fumigation and the release of soil nitrogen: a rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biology and Biochemistry, 17(6): 837-842.
Cadisch, G.; Handayanto, E.; Malama, C.; Seyni, F.; Giller, K.E. 1998. N recovery from legume prunings and priming effects are governed by the residue quality. Plant and Soil, 205: 125-134.
Constantinides, M.; Fownes, J.H. 1994. Nitrogen mineralization from leaves and litter of tropical plants: relationship to nitrogen, lignin and soluble polyphenol concentrations. Soil Biology and Biochemistry, 26(1): 49-55.
EMBRAPA-CPAA. 1997. Dinâmica do solo, da vegetação e efeitos ambientais sob sistemas agroflorestais em pastagens degradadas. Relatório anual sub-projeto 1. EMBRAPA-CPAA, Manaus, Brazil.
FAO-UNESCO. 1990. Soil map of the world, revised legend. Food and Agriculture Organization of the United Nations, Rome.
Fernandes, E.C.M.; Perin, R.; Wandelli, E.; Souza, S.G. de; Matos, J.C.; Arco Verde, M.; Ludewigs, T.; Neves, A. 1999 Agroforestry systems to rehabilitate abandoned pastoreland in the brazilian amazon. In: Jiménez, F.; Beer, J. (eds.). Proceedings International Symposium on Multi-Strata Agroforestry Systems with Perennial Crops,Turrialba (Costa Rica)22-27 Feb 1999, p. 24-26.Giller, K.E. 2001. Nitrogen fixing in tropical cropping systems. 2nd ed. CABI Publishing, New York. 423pp.
Haggar, J.P.; Tanner, E.V.J.; Beer, J.W.; Kass, D.C.L. 1993. Nitrogen dynamics of tropical agroforestry and annual cropping systems. Soil Biology & Biochemistry, 25(10): 1363-1378.
Handayanto, E.; Giller, K.E.; Cadisch, G.. 1997. Regulating N release from legume tree prunings by mixing residues of different quality. Soil Biology and Biochemistry, 29(9/10): 1417-1426.
Henriksen, T.; Breland, T. 2002. Carbon mineralization, fungal and bacterial growth, and enzyme activities as affected by contact between crop residues and soil. Biology and Fertility of Soils, 35: 41-48.
Hofer, H.; Hanagarth, W.; Garcia, M.; Martius, C.; Franklin, E.; Rombke, J.; Beck, L. 2001. Structure and function of soil fauna communities in Amazonian anthropogenic and natural ecosystems. European Journal of Soil Biology, 37(4): 229-235.
Isaac, L.; Wood, C.W.; Shannon, D.A. 2003. Pruning management effects on soil carbon and nitrogen in contour-hedgerow cropping with Leucaena leucocephala (Lam.) De Wit on sloping land in Haiti. Nutrient Cycling in Agroecosystems, 65(3): 253-263.
Kang, B.T.; Wilson, G.F.; Sipkens, L. 1981. Alley cropping maize (Zea-mays-L) and leucaena (Leucaena- leucocephala Lam) in Southern Nigeria. Plant and Soil, 63(2): 165-179.
Lehmann, J.; Schroth, G.; Zech, W. 1995. Decomposition and nutrient release from leaves, twigs, and roots of three alley-cropped tree legumes in Central Togo. Agroforestry Systems, 29: 21-36.
Lehmann, J.; Muraoka, T.; Zech, W. 2001. Root activity patterns in an Amazonian agroforest with fruit trees determined by 32P, 33P, and 15N applications. Agroforestry Systems, 52: 185-197.
Mafongoya, P.L.; Nair, P.K.R.; Dzowela, B.H. 1998a. Mineralization of nitrogen from decomposing leaves of multipurpose trees as affected by their chemical composition. Biology and Fertility of Soils, 27: 143-148.
Mafongoya, P.L.; Giller, K.E.; Palm, C.A. 1998b. Decomposition and nitrogen release patterns of tree prunings and litter. Agroforestry Systems, 38(1-3): 77-97.
McGrath, D.; Comerford, N.; Duryea, M. 2000. Litter dynamics and monthly fluctuations in soil phosphorus availability in an Amazonian agroforest. Forest Ecology and Management, 131: 167-181.
Minitab Statistical Software. 2000. Release 13.1.
Mulongoy, K.; Ibewiro, E.B.; Oseni, O.; Kilumba, N.; Opara-Nadi, A.O.; Osonubi, O. 1993. Effect of management practices on alley-cropped maize utilization of nitrogen derived from prunings on a degraded Alfisol in south-western Nigeria, In: Mulongoy, K.; and Merckx, R. (Eds). Soil Organic Matter Dynamics and Sustainability of Tropical Agriculture. John Wiley and Sons, Chichester. p. 223-230.
Palm, C.A. 1995. Contribution of agroforestry trees to nutrient requirements of intercropped plants. Agroforestry Systems, 30: 105-124.
Palm, C.A.; Sanchez, P.A. 1990. Decomposition and Nutrient Release Patterns of the Leaves of Three Tropical Legumes. Biotropica, 22(4): 330-338.
Palm, C.A.; Sanchez, P.A. 1991. Nitrogen release from the leaves of some tropical legumes as affected by their lignin and polyphenolic contents. Soil Biology and Biochemistry, 23(1): 83-88.
Russo, R.O.; Budowski, G. 1986. Effect of Pollarding Frequency On Biomass of Erythrina- Poeppigiana As a Coffee Shade Tree. Agroforestry Systems, 4(2): 145-162.
Sanchez, P.A.; Bandy, D.E.; Villachica, J.H.; Nicholaides, J.J. 1982. Amazon Basin soils: management for continuous crop production. Science, 216(4548): 821-827.
Schroth, G., Silva, L.F. da; Seixas, R.; Teixeira, W.G.; Macedo, J.L.V.; Zech, W. 1999. Subsoil accumulation of mineral nitrogen under polyculture and monoculture plantations, fallow and primary forest in a ferralitic Amazonian upland soil. Agriculture Ecosystems & Environment, 75(1-2): 109-120.
Silva, F.C.d. 1999. Manual de análises químicas de solos, plantas, e fertilizantes Embrapa Communicação para Transferência de Tecnologia, Brasilia, DF. 370pp.
Tian, G.; Kang, B.T.; Brussard, L. 1992. Biological effects of plant residues with contrasting chemical compositions under humic tropical conditions - decomposition and nutrient release. Soil Biology and Biochemistry, 24(10): 1051-1060.
Uguen, K. 2001. Influence des arbres sur la matière organique et l’azote du sol dans un système agroforestier en Amazonie centrale. Doctoral Dissertation, Institut Nacional Agronomique Paris-Grignon, Paris. 158pp.
U.S. Soil Survey Staff. 1998. Keys to soil taxonomy. 8th ed. U.S. Department of Agriculture, Natural Resources Conservation Service, Washington, D. C.
Vohland, K.; Schroth, G. 1999. Distribution patterns of the litter macrofauna in agroforestry and monoculture plantations in central Amazonia as affected by plant species and management. Applied Soil Ecology, 13(1): 57-68.
Wilson, G.F.; Kang, B.T.; Mulongoy, K. 1986. Alley cropping - trees as sources of green-manure and mulch in the tropics. Biological Agriculture & Horticulture, 3(2-3): 251-267.
Recebido em 17/06/03
Aceito em 26/02/07