Tree communities of white-sand and terra-firme forests of the upper Rio Negro
Comunidades de árvores em florestas de campinarana e de terra-firme do alto Rio Negro
Juliana STROPP1, Peter VAN DER SLEEN2, Paulo Apóstolo ASSUNÇÃO3, Adeilson Lopes da SILVA4, Hans TER STEEGE5
1 Ecology and Biodiversity Group, Utrecht University; justropp@gmail.com
2 Forest Ecology and Management Group, Wageningen University; peter.vandersleen@wur.nl
3 Autonoumos; apostolo.paulo@yahoo.com.br
4 Instituto Socioambiental – Programa Rio Negro; adeilson@socioambiental.org
5 Ecology and Biodiversity Group, Utrecht University; h.tersteege@uu.nl
Abstract
The high tree diversity and vast extent of Amazonian forests challenge our understanding of how tree species abundance and composition varies across this region. Information about these parameters, usually obtained from tree inventories plots, is essential for revealing patterns of tree diversity. Numerous tree inventories plots have been established in Amazonia, yet, tree species composition and diversity of white-sand and terra-firme forests of the upper Rio Negro still remain poorly understood. Here, we present data from eight new one-hectare tree inventories plots established in the upper Rio Negro; four of which were located in white-sand forests and four in terra-firme forests. Overall, we registered 4703 trees ≥ 10 cm of diameter at breast height. These trees belong to 49 families, 215 genera, and 603 species. We found that tree communities of terra-firme and white-sand forests in the upper Rio Negro significantly differ from each other in their species composition. Tree communities of white-sand forests show a higher floristic similarity and lower diversity than those of terra-firme forests. We argue that mechanisms driving differences between tree communities of white-sand and terra-firme forests are related to habitat size, which ultimately influences large-scale and long-term evolutionary processes.
Keywords: tree communities, tree inventory plots, terra-firme forest, white-sand forest, upper Rio Negro
Resumo
A vasta extensão e a alta diversidade de árvores das florestas na Amazônia desafiam a nossa compreensão sobre como variam a composição e abundância de espécies arbóreas ao longo desta região. Informações sobre estes parâmetros, geralmente obtidas a partir de inventários florísticos, são fundamentais para análises sobre padrões de diversidade. Embora inúmeros inventários florísticos tenham sido estabelecidos na Amazônia, a composição de espécies e a diversidade de árvores das florestas de campinarana e de terra-firme, na região do alto Rio Negro, permanecem ainda pouco conhecidas. Aqui, apresentamos dados de inventários florísticos de oito parcelas de um hectare estabelecidas em florestas de campinarana e de terra-firme do alto Rio Negro. Ao todo, registramos 4.703 indivíduos com diâmetro à altura do peito ≥ 10 cm pertencentes a 49 famílias, 215 gêneros e 603 espécies. As comunidades de árvores das florestas de campinarana e terra-firme do alto Rio Negro diferem entre si, sendo que as florestas de campinaranas são mais similares e menos diversas que as florestas de terra-firme. Sugerimos que os mecanismos que causam diferenças entre as comunidades arbóreas das florestas de campinarana e terra-firme são principalmente relacionados a diferenças do tamanho do habitat, isto porque o tamanho da área do habitat influencia processos evolutivos que ocorrem em grande escala temporal e espacial.
Palavras-chave: comunidades de árvores, parcelas de inventário florestal, floresta de terra-firme, florestas de campinarana, alto Rio Negro.
Introduction
The Amazon basin contains 40% of the world’s remaining tropical rainforests and may harbor approximately 12,500 tree species (Hubbell et al. 2008). Attempts to systematically analyze tree species diversity and forest structure of Amazonian forests date back to 1930 (Pires et al. 1953; Davis and Richards 1934). Over the past six decades (oldest plots from the 30ies), a total of about 900 one-hectare tree inventory plots has been established (Amazon Tree Diversity Network ATDN, 2010). For the upper Rio Negro region, tree inventory plots are particularly scarce and thus under-represented in the ATDN dataset. This shortcoming is relevant because unlike prevailing landscapes in Amazonia, where terra-firme forests are the dominant forest type, the upper Rio Negro harbors extensive areas of white-sand forests, which covers ca. 5-10% of the total area of Amazonia (IBGE 1997).
White-sand forests are the dominant vegetation on non-flooded sandy soils in the upper Rio Negro regions. These forests are also classified as ‘campinarana’, ‘campinarana arbustiva’, or ‘campinarana florestada’ (IBGE, 1997) and are commonly referred as ‘campinarana’ (Brazil), ‘varillal’ (Colombia), ‘wallaba’ or ‘white-sand forests’ (Guyana) (Anderson 1981; Banki 2010). White-sand forests in the upper Rio Negro region are characterized by Eperua leucantha (Fabaceae), Hevea spp. and Micrandra sprucei (Euphorbiaceae) (Dezzeo et al. 2000; Boubli 2002; Abraao et al. 2008). In contrast, terra-firme forests grow on non-flooded clayey soils (oxisols and latosols); they are also classified as ‘floresta ombrófila’ (IBGE 1997). Previous tree inventories conducted in the upper Rio Negro region show that terra-firme forests are typically characterized by Protium spp. (Burseraceae), Caryocar spp. (Caryocaraceae), Licania spp. (Chrysobalanaceae), Swartzia spp. (Fabaceae), Eschweilera spp. (Lecythydaceae) and Vochysia spp. (Vochysiaceae) (Dezzeo et al. 2000; Boubli 2002). The classification of white-sand forests and terra-firme forests are broad and multiple, somehow imprecise, definition of these two forest types. Differences in forest structure, tree species diversity, and composition within each of these categories may arise from local environmental conditions as well as from random factors, such as, e.g., dispersal limitation, and occurrence of pathogens.
Tree communities of white-sand and terra-firme forests vary in their species diversity and composition (Dezzeo et al. 2000; ter Steege et al. 2000; Boubli 2002; Banki 2010). Tree communities of white-sand forests have a lower diversity and a distinctively different floristic composition in comparison to tree communities of terra-firme forests (ibid.). The lower diversity in white-sand forests has been explained principally by 1) the indirect effects of soil nutrient availability on species diversity, e.g., low ecosystem dynamics, (ter Steege and Hammond 2001), 2) ‘density-dependence’ effects (Janzen, 1974), as well as 3) the patchy configuration and the small size of the area in which white-sand forests occur (ter Steege et al. 2000). Each of these explanations emphasizes different mechanisms regulating tree species diversity of white-sand forests. In addition, variation in tree species composition has been attributed to the direct effect of physical environmental characteristics in these two forest types: white-sand and terra-firme forests differ in the availability of light in the understory (Coomes and Grubb 1996), soil water (Luizão et al. 2007), as well as in edaphic features, especially H+ (high in white-sand) and Al toxicity (high in terra-firme) (Luizão et al. 2007). As a consequence of different physical environmental conditions, tree species have become highly specialized in their habitat, and therefore may suffer a disadvantage when establishing in non-typical habitat. Moreover, an interaction between soil nutrient availability and herbivory may constrain species distribution in their typical habitat (Janzen 1974; Fine et al. 2004).
Differences in species composition and diversity between tree communities of white-sand and terra-firme forests suggest that these communities may have a relatively independent evolutionary history. The marked difference in the habitat size of these two forest types may have influenced evolutionary history and, therefore, the current tree diversity of white-sand and terra-firme forest (ter Steege et al. 2000). This is because habitat size determines the number of species a habitat can support: large habitats typically host more species, and potentially, more co-generic species than do smaller ones (Rosenzweig 1995; Harnik et al. 2010). Variation in habitat size could, therefore, generate differences in species diversity as well as in species composition.
Here, we present data of eight new one-hectare tree inventory plots located in the white-sand and terra-firme forests of the upper Rio Negro. We aim at comparing the forest structure, tree species diversity, and tree species composition among these plots. We discuss our findings focusing, particularly, on the contribution of habitat size to differences in tree species diversity and composition of white-sand and terra-firme forests.
Material and Methods
STUDY AREA
The study was carried out in Brazil in the upper Rio Negro region which is located at the south-western border of the Guiana shield (Figure 1). The region has an annual rainfall ranging from 2500 to 3000 mm and is one of the wettest and least seasonal regions in Amazonia (Sombroek 2001). The average yearly temperature is 24 oC (Bezerra et al. 1976). The upper Rio Negro region belongs to the Rio Negro-Juruena geo-chronological province (Kroonenberg and de Roever 2010). Its bedrocks are mainly classified as granites and gneisses, which originated during the Precambrian (Bezerra et al. 1976). The relief in this region is influenced mainly by differences in the weathering resistance of the bedrocks (Kroonenberg and de Roever 2010). In hilly areas, the dominant soils are well-drained clayey latosols and oxisols (Dubroeucq and Volkoff 1998; Bezerra et al. 1976). By contrast, in the widespread flat areas, sandy podzols occur, the origin of which can be attributed to in-situ weathering, alluvial deposition, or podzolization (Anderson 1981; Quesada 2008).
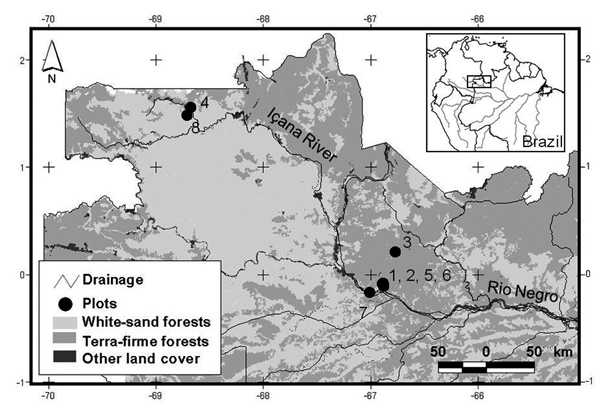
Figure 1 - Locations of the eight one-hectare tree inventory plots established in the upper Rio Negro (black dots). Plots 1, 2, 3, and 4 were placed in terra-firme forests and plots 5, 6, 7 and 8 were placed in white-sand forests. Plot 7 is located nearby the city of São Gabriel da Cachoeira. Site names and plots’ geographical coordinates are given in Table 1. The base map is a courtesy of Instituto Socioambiental (ISA).
Table 1 - Total number of trees, total number of species, and Fisher’s alpha index of eight one-hectare plots established in the upper Rio Negro. TF indicates terra-firme plots; WS indicates white-sand plots; SGC indicates plots established nearby the city of São Gabriel da Cachoeira; Içana indicates plots established at the middle Içana River
Plot No. (Forest type) |
Site name |
latitude |
longitude |
No. of trees |
No. of species |
Fisher’s alpha |
Plot dimensions |
1 (TF) |
COMAR-01 (SGC) |
-0.084 |
-66.888 |
523 |
126 |
52.69 |
100 x 100 |
2 (TF) |
COMAR-02 (SGC) |
-0.086 |
-66.891 |
537 |
142 |
63 |
60 x 140 + 40x40 |
3 (TF) |
Pico da Neblina National Park (SGC) |
0.211 |
-66.771 |
505 |
155 |
76.35 |
40 x 250 |
4 (TF) |
Igarapé Pamaáli (Içana) |
1.553 |
-68.676 |
614 |
192 |
95.92 |
40 x 250 |
5 (WS) |
COMAR-03 (SGC) |
-0.101 |
-66.88 |
547 |
102 |
36.95 |
100 x 100 |
6 (WS) |
COMAR-04 (SGC) |
-0.111 |
-66.881 |
642 |
95 |
30.8 |
100 x 100 |
7 (WS) |
Itacoatiara-Mirim (SGC) |
-0.166 |
-67.011 |
611 |
87 |
27.73 |
40 x 250 |
8 (WS) |
Jandú-Cachoeira (Içana) |
1.481 |
-68.711 |
724 |
116 |
39.01 |
40 x 250 |
1 Plots had a square or rectangular shape and were divided in 25 subplots of 20 x 20 m; one exception presents Plot 2, for which we had to set up two adjacent plots of 60 x 140 m and 40 x 40 m each in order to assure similar edaphic conditions.
TREE INVENTORY PLOTS AND BOTANICAL IDENTIFICATION
We established eight one-hectare tree inventory plots, four of which were placed in mature white-sand forests and four in terra-firme forests. Two plots were set up on indigenous land, in the ‘Terra Indígena do alto Rio Negro’, at the middle Içana River. The remaining six plots were set up in the vicinity of São Gabriel da Cachoeira: four at the air force area ‘Comando Aéreo Regional – COMAR’, one at Pico da Neblina National Park, and one at the indigenous community of ‘Itacoatiara-Mirim’ (Figure 1, Table 1). To set up the plots, we collaborated with the ‘Escola Indígena Baniwa Coripaco – Pamaáli’ (EIBC – Pamaáli) and with the ‘Instituto Federal de Educação, Ciência e Tecnologia do Amazonas (Ifam), campi São Gabriel da Cachoeira and Tunuí)’.
In each plot, we measured the diameter of all trees ≥ 10 cm at breast height, i.e., DBH at 1.3 m. We tagged trees of all morpho-species and collected at least one sample per morpho-species. The fertile botanical collections were deposited at the herbarium of the ‘Instituto Nacional de Pesquisas da Amazônia’ (INPA, Manaus) and at the herbarium of Ifam (campus São Gabriel da Cachoeira). Sterile material was deposited at the herbarium of the ‘Instituto Federal de Educação, Ciência e Tecnologia do Amazonas’ (EAF-Manaus). All tree species were identified at the INPA herbarium. Sterile material of the families Annonaceae, Moraceae, and Urticaceae was identified by P. J. M. Maas and C. C. Berg. In the field, we collected at least one sample per morpho-species. The identification procedure revealed in a few cases that one morpho-species assigned to multiple trees during the field work, turned out to represent in fact two different species. To account for these cases, we created a ‘complex of species’ which reports the two species names assigned in the herbarium.
DATA ANALYSIS
Forest structure
We tested whether white-sand and terra-firme plots differ in the diameter size-class distributions by performing a Kolmorogov-Smirnov test. To this end, we first classified trees according to their DBH in 9 classes at increments of 5 cm (i.e., 10 - < 15 cm, …, 45 - < 55 cm) and one including all trees with DBH ≥ 55 cm. Afterwards, we calculated the total abundance of each DBH size-class. We, furthermore, calculated total basal area and stem density for each plot.
Tree species composition and diversity
Hereafter we refer to the trees sampled in the four white-sand plots as tree community of white-sand forests and to the trees sampled in the four terra-firme plots as tree community of terra-firme forests. We established relative species abundance curves for the tree communities of white-sand and terra-firme forests. We tested differences in tree species composition among these two tree communities with a multi-response permutation procedure (MRPP) (McCune et al. 2002). For this procedure, we used the Bray-Curtis Index as a measure of floristic dissimilarity (FD) and we defined forest type (i.e., white-sand and terra-firme forests) as group variables. We assessed the variation in tree species composition by performing a 2 dimensions non-metric multidimensional scaling (NMDS) (McCune et al. 2002). We performed NMDS because we sampled tree communities in a discontinued manner (i.e., plots placed either in white-sand or terra-firme forests). For the NMDS, we first calculated the relative density of each species in each plot by dividing the absolute abundance of species i in plot p by the total absolute abundance of trees in plot p. We used relative density in the NMDS ordination (see Costa et al. 2009). We used the Bray-Curtis Index as a measure of floristic dissimilarity and 5000 runs for randomized data, as well as for real data. We calculated the percentage of variation explained by the two NMDS axes using the same Bray-Curtis Index as used for the ordination. We determined whether family, genus, and species were indicators of either white-sand forests or terra-firme forests by performing an indicator species analysis (ISA). This analysis calculates an indicator index for each family, genus, and species. This indicator index is derived from the total abundance of each family, genera, and species registered in each plot. This index is maximum when the respective taxa is present in all plots of one forest type and absent in all plots of the other forest type (Dufrene and Legendre 1997). Finally, we estimated tree species diversity for each plot by calculating Fisher’s alpha (Fisher et al. 1943). We applied the Fisher’s alpha index because it is relatively insensitive to tails of the distribution, i.e., to differences in the number of rare or very abundant species (Magurran 1988). We performed a single factor analysis of variance (ANOVA) to test whether tree diversity varies among white-sand and terra-firme plots. For ANOVA, we used SPSS v.16. We used PC-ORD version 5.0 for performing MRPP, NMDS, and ISA analyses (McCune and Mefford, 1999). The Kolmorogov-Smirnov was performed in the R statistical and programming environment (Ihaka and Gentleman 1996).
Results
FOREST STRUCTURE
The total basal area of the eight plots varied between 19.4 m2 and 31.4 m2. Interestingly, both maximum and minimum total basal area values were registered for white-sand plots: plot 8 (Jandú-Cachoeira) and plot 7 (Itacoatiara-Mirim), respectively. For terra-firme plots, total basal area ranged from 27.4 m2 (plot 4, Igarapé Pamaáli) to 30.40 m2 (plot 1, COMAR-01). For all plots, Eperua leucantha had the highest total basal area, followed by Eperua purpurea and Monopteryx uaucu. On white-sand plots, the three most abundant species also had the largest total basal area (E. leucantha, E. purpurea, Aldina heterophylla). By contrast, on terra-firme plots, M. uaucu, had the highest total basal area, although it was only the third abundant species. Additionally, Micrandra spruceana had the second highest total basal area (fifth in abundance) and Clathrotropis macrocarpa the third in total basal area. Stem density varied between 547 and 724 trees ha-1 in white-sand plots and between 505 and 614 trees ha-1 in terra-firme plots. The diameter size-class distributions were similar for white-sand and terra-firme plots (D = 0.3; p = 0.7) (Figure 2).
Overall, 41% of trees had a DBH between 10 and 15 cm. On all white-sand plots, the density of these small trees ranged from 191 trees ha-1 (34.91%) to 392 trees ha-1 (54.14%). For terra-firme plots, this density varied between 195 trees ha-1 (38.61%) and 292 trees ha-1 (47.39%).
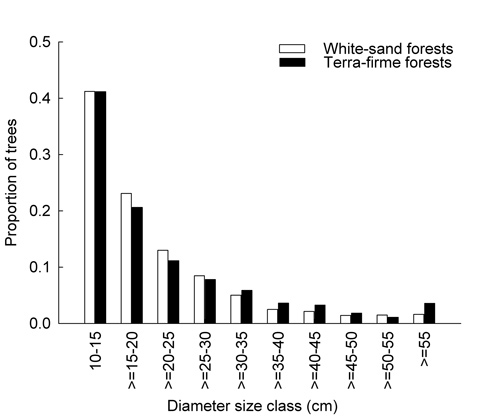
Figure 2 - Diameter size-class distributions (measured in centimeters of diameter at breast height) in eight one-hectare plots in the upper Rio Negro. Proportions are based on 2,524 tree sampled in white-sand plots and a 2,179 trees sampled in terra-firme plots.
TREE SPECIES COMPOSITION AND DIVERSITY
We registered 4,703 trees ≥ 10 cm DBH, belonging to 49 families, 215 genera, and 603 species in eight one-ha plots. In white-sand plots, we found 2,524 trees, 39 families, 124 genera, and 290 species (Table 1). In terra-firme plots, we found 2,179 trees, 44 families and 183 genera, and 417 species. These inventories include potentially one new observation for Brazil (Ephedranthus guianensis) (Maas et al., 2010), a first observation for the upper Rio Negro region (Pseudoxandra duckei – P. J. M. Maas, pers. comm.), and an undescribed species (Batocarpus sp. nov. – C. C. Berg, pers. comm.). Twenty two individual trees (0.47% of all trees) were not identified at the family level. Additionally, 680 trees (14.45%), accounting for 142 morpho-species were only identified at the genus level, and 204 trees (4.33%), representing 64 morpho-species, remain to be confirmed (Appendix). For 112 trees belonging to 6 morpho-species we assigned the category ‘complex of species’ (see Methods).
Overall, the most abundant family identified in white-sand and terra-firme forests was Fabaceae with 2,222 trees (47%), followed by Sapotaceae with 288 trees (6%) and Myristicaceae with 283 trees (6%). Fabaceae was the most abundant family in both white-sand and terra-firme plots, accounting for 1,451 trees (57%) and 771 trees (35%), respectively. In white-sand plots, the second and third most abundant families were Sapotaceae with 175 trees (7%) and Euphorbiaceae with 102 trees (4%), whereas in terra-firme plots, the second and third most abundant families were Myristicaceae with 220 trees (10%) and Euphorbiaceae with 165 trees (8%) (Figure 3). The most abundant species in white-sand plots was Eperua leucantha with 681 trees (26%), followed by Eperua purpurea with 191 trees (8%), and Aldina heterophylla with 137 trees (5%). Clathrotropis macrocarpa was the most abundant species in terra-firme plots with 138 trees (6%), followed by Pterocarpus sp. with 126 trees (6%), and Monopteryx uaucu with 74 trees (3%). The range of relative abundance observed for the four most abundant species in white-sand plots is larger (0.05 – 0.26) than the same range observed for terra-firme-plots (0.03 – 0.06) (see Table 2 and Figure 4).
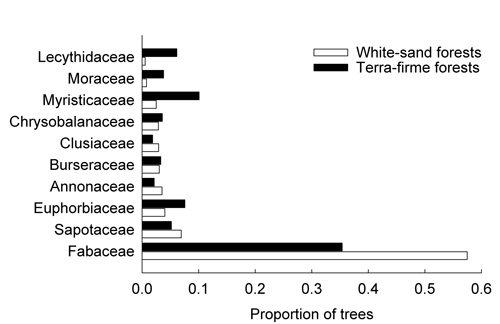
Figure 3 - Proportion of trees of the ten most abundant families encountered in the eight one-hectare tree inventory plots. Proportion of trees is based on 2,524 trees encountered in four one-hectare white-sand plots and 2,179 trees registered in four one-hectare terra-firme plots.
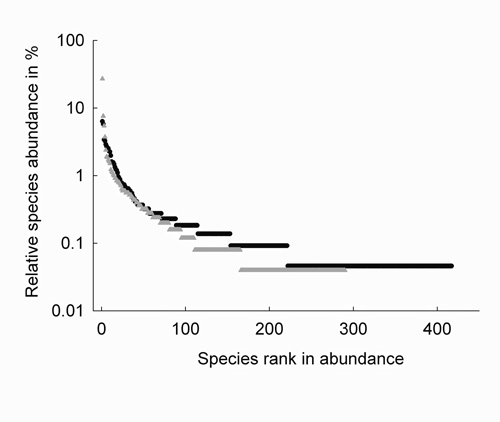
Figure 4 - Relative species abundances in the four one-ha tree inventory plots established in white-sand forests (▲) and terra-firme forests (▲). Total number of trees (DBH ≥ 10 cm) encountered in white-sand and terra-firme plots is 2,524 and 2,179, respectively. The total number of species registered in white-sand and terra-firme plots are 290 and 417.
Table 2 - Number of trees and relative abundance (in parenthesis) of the ten most abundant species encountered in four one-ha plots established in terra-firme forests (plots 1, 2, 3, and 4) and in white-sand forests (plots 5, 6, 7, and 8); plot codes and locations are given in Table 1
Family |
Species |
plot 1 |
plot 2 |
plot 3 |
plot 4 |
Euphorbiaceae |
Micrandra spruceana (Baill.) R.E. Shult. |
20 (0.04) |
15 (0.03) |
0 (0.00) |
36 (0.07) |
Fabaceae |
Andira unifloliolata Ducke |
9 (0.02) |
20 (0.04) |
0 (0.00) |
34 (0.07) |
Fabaceae |
Clathrotropis macrocarpa Ducke |
28 (0.05) |
47 (0.09) |
21 (0.03) |
42 (0.08) |
Fabaceae |
Monopteryx uaucu Spruce ex Benth. |
59 (0.11) |
0 (0.00) |
15 (0.02) |
0 (0.00) |
Fabaceae |
Pterocarpus sp.2 INPA |
63 (0.12) |
42 (0.08) |
0 (0.00) |
21 (0.04) |
Fabaceae |
Swartzia polyphylla DC. |
15 (0.03) |
22 (0.04) |
0 (0.00) |
19 (0.04) |
Fabaceae |
Swartzia tomentifera (Ducke) Ducke |
2 (0.00) |
4 (0.01) |
30 (0.05) |
23 (0.05) |
Lecythidaceae |
Eschweilera pedicellata (Rich.) S. A. Mori |
24 (0.05) |
19 (0.04) |
0 (0.00) |
12 (0.02) |
Myristicaceae |
Iryanthera ulei Warb. |
7 (0.01) |
18 (0.03) |
9 (0.01) |
15 (0.03) |
Myristicaceae |
Virola calophylla/pavonis |
7 (0.01) |
7 (0.01) |
35 (0.06) |
1 (0.00) |
|
plot 5 |
plot 6 |
plot 7 |
plot 8 |
|
Fabaceae |
Eperua leucantha Benth. |
212 (0.39) |
282 (0.44) |
159 (0.26) |
28 (0.04) |
Fabaceae |
Eperua purpurea Benth. |
51 (0.09) |
0 (0.00) |
108 (0.18) |
32 (0.04) |
Fabaceae |
Aldina heterophylla Spruce ex Benth. |
33 (0.06) |
25 (0.04) |
44 (0.07) |
35 (0.05) |
Fabaceae |
Inga sp.1 |
0 (0.00) |
0 (0.00) |
0 (0.00) |
94 (0.13) |
Fabaceae |
Monopteryx uaucu Spruce ex Benth. |
12 (0.02) |
25 (0.04) |
23 (0.04) |
0 (0.00) |
Fabaceae |
Taralea oppositifolia Aubl. |
11 (0.02) |
12 (0.02) |
16 (0.03) |
9 (0.01) |
Sapotaceae |
Pradosia cf. mutisii Cronquist |
2 (0.00) |
10 (0.02) |
12 (0.02) |
23 (0.03) |
Malvaceae |
Scleronema micranthum (Ducke) Ducke |
9 (0.02) |
3 (0.00) |
30 (0.05) |
1 (0.00) |
Euphorbiaceae |
Micrandra sprucei (Müll. Arg.) R.E. Schult. |
0 (0.00) |
0 (0.00) |
0 (0.00) |
41 (0.06) |
Clusiaceae |
Haploclathra paniculata (Müll. Arg.) R.E. Schult. |
0 (0.00) |
10 (0.02) |
0 (0.00) |
28 (0.04) |
Floristic composition in white-sand plots was significantly different from terra-firme (MRPP: A = 0.16; p = 0.007). The floristic dissimilarity ranged from 0.38 to 1 among the pairs of eight plots, from 0.38 to 0.84 among pairs of white-sand plots, and from 0.48 to 0.87 among pairs of terra-firme plots. Considering pairs of white-sand and terra-firme plots, the floristic dissimilarity ranged from 0.81 to 1. The species indicator analysis revealed that there are fewer significant indicator families, genera, and species in white-sand (3 families, 8 genera, and 5 species) than in terra-firme forests (5 families, 12 genera, 9 species) (Table 3 and 4). Annonaceae, Apocynaceae, and Fabaceae were indicator families for white-sand forests. Lecythidaceae, Meliaceae, Moraceae, Myristicaceae, and Quiinaceae were indicator families for terra-firme forests (Table 3). Interestingly, Fabaceae, which represents the most abundant family in both forest types and which is an indicator family for white-sand forests, had a higher number of species in terra-firme forests than in white-sand forests.
Table 3 - Number of trees belonging to the families, which are significantly associated with one of the two forest types (p < 0.05); (*) indicates the forest type which the families are associated with
Family |
Number of trees in white-sand |
Number of trees in terra-firme |
Annonaceae |
89* |
47 |
Apocynaceae |
46* |
12 |
Fabaceae |
1451* |
771 |
Lecythidaceae |
14 |
134* |
Meliaceae |
1 |
10* |
Moraceae |
20 |
83* |
Myristicaceae |
63 |
220* |
Quiinaceae |
0 |
6* |
Table 4 - Number of trees belonging to the genera, which are significantly associated with one of the two forest types (p < 0.05); (*) indicates the forest type which the genera are associated with
Family |
Genus |
Number of trees in white-sand |
Number of trees in terra-firme |
Annonaceae |
Bocageopsis |
0 |
17* |
Fusaea |
0 |
8* |
|
Xylopia |
67* |
4 |
|
Araceae |
Dendropanax |
7* |
0 |
Clusiaceae |
Symphonia |
1 |
7* |
Euphorbiaceae |
Sandwithia |
0 |
43* |
Fabaceae |
Clathrotropis |
15 |
138* |
Eperua |
872* |
50 |
|
Macrolobium |
80* |
5 |
|
Swartzia |
45 |
136* |
|
Taralea |
48* |
6 |
|
Lecythidaceae |
Couratari |
0 |
8* |
Eschweilera |
14 |
112* |
|
Moraceae |
Brosimum |
18 |
44* |
Myristicaceae |
Virola |
6 |
90* |
Rubiaceae |
Chimarrhis |
0 |
15* |
Pagamea |
30* |
1 |
|
Sapotaceae |
Manilkara |
42* |
3 |
Pradosia |
52* |
3 |
|
Vochysiaceae |
Erisma |
1 |
38* |
Four-hundred-one species occurred in only one of the eight plots; out of these 401 species, 154 were registered in only one of the white-sand plots and 247 in only one of the terra-firme plots. Two-hundred-sixty-two species occurred with only one individual. Out of this total, 92 species were encountered in white-sand forests and 170 species in terra-firme forests.
The two axes of the NMDS captured 77% of the floristic variation. The first and the second axis explained 61% and 16% of the floristic variation among the eight one-ha plots, respectively. The variation on the first axis can be explained by the abundance of different species present in each forest type: plots placed in terra-firme forests are located on the left side of Figure 5, plots in white-sand forests on the right. The second axis is related to geographic distance: plots established close to São Gabriel da Cachoeira are clustered at the upper part of Figure 5 and plots established at middle Içana River are found at the lower part.
Tree diversity, per hectare, measured here by Fisher’s alpha, ranged from 27.3 (white-sand plots) to 95.9 (terra-firme plots, Table 1). For white-sand plots, tree diversity ranged from 27.7 (plot 7, Itacoatiara-Mirim) to 39.0 (plot 8, Jandú-Cachoeira), respectively. For terra-firme plots, tree diversity ranged from 52.7 (plot 1, COMAR-1) to 95.9 (plot 4, Igarapé Pamaáli), respectively. Tree diversity was on average significantly lower in white-sand plots than in terra-firme plots (F = 15.66; p = 0.007).
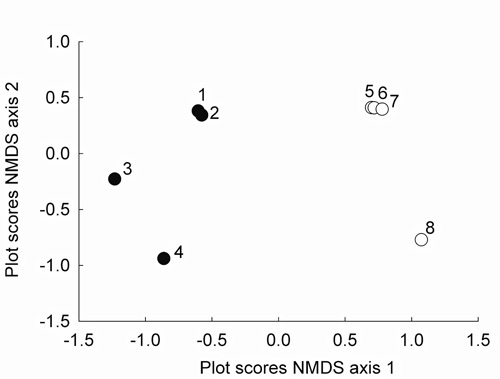
Figure 5 - Ordination diagram of non-metric multidimensional scaling (NMDS) of the eight one-hectare tree inventory plots. The first and the second axis explained 61% and 16% of the floristic variation, respectively. Terra-firme plots are represented by black dots and white-sand plots by white dots. The locations of the plots are given in Figure 1 and Table 1.
Discussion
OVERVIEW
Tree communities of terra-firme and white-sand forests of the upper Rio Negro show a significant floristic dissimilarity. Plots established in white-sand forests were more similar to each other and less diverse than plots established in terra-firme forests. This dissimilarity is to a large extent caused by 1) differences in relative abundance of highly abundant species, 2) the occurrence of families, genera, and species which are restricted to either of the two forest types, and 3) the low number of rare species registered for white-sand forests.
FOREST STRUCTURE
The diameter size-class distributions observed here are comparable to those of other tree inventory plots established in the upper Rio Negro (Dezzeo et al. 2000; Boubli 2002). Similar to Boubli (2002), we found that 40% of the trees registered in white-sand and terra-firme plots have a DBH between 10 and 15 cm. Comparable diameter size-class distributions have been commonly reported for terra-firme forests in other regions of Amazonia (Amaral et al. 2000; Lima Filho et al. 2001; Pitman et al. 2002; Oliveira and Amaral 2004; Oliveira et al. 2008).
TREE SPECIES COMPOSITION
Patches of white-sand forests are typically surrounded by terra-firme forests in Amazonia. Seed dispersal between these two forests types may occur regularly, as indicated by some tree species, which do occur in both forest types. These tree species, however, generally show high differences in abundance (see Appendix). We, therefore suggest that seed dispersal may be of minor importance for limiting species occurrence in either forest type. Arguably, a main limitation to species establishment occurs at the seedling stage. In fact, tree seedling mortality seems to be higher when seedlings attempt to establish in their non-typical habitat (Stropp et al. 2011), indicating a habitat specificity that is likely to be related to the edaphic characteristics of these two forest types.
Several mechanisms have been proposed to explain habitat specificity of trees in white-sand and terra-firme forests. The hypothesis of a trade-off between plant investment in growth and herbivore defenses proposes that tree species, which typically grow in nutrient-limited conditions, such as white-sand species, allocate more of their resources in herbivore defense and as a consequence have to allocate less in growth (Fine et al. 2004). Conversely, species occurring in less nutrient-limited conditions, such as terra-firme tree species, invest their resources in growth and relatively less in herbivore defenses. This divergence in the growth-defense strategy may be a result of long-term habitat specialization (Fine et al. 2004; Fine et al. 2006). This trade-off between growth and defense drives habitat specialization of trees in white-sand and terra-firme in western Amazonia (Fine et al. 2004) but seems to be of minor importance for habitat specialization in these two forest types of the upper Rio Negro. Stropp et al. (2011) have shown that seedlings of white-sand and terra-firme habitat specialist species, including the highly abundant E. purpurea and C. macrocarpa, suffer similar herbivory rates and show similar growth performance, regardless of the forest type in which they grow. An explanation for these diverging findings might be provided by additional soil characteristics: in the upper Rio Negro soil characteristics may have a more direct limiting effect on seedling establishment and growth. An experiment conducted in French Guiana (Guiana shield), has shown that tree seedlings growth of species typically occurring either in white-sand or terra-firme forests is not limited by phosphate, but seems to respond differently to soil drought (Baraloto et al. 2006). Although H+ and Al toxicity have been shown to limit species distribution between white-sand and terra-firme forests in central Amazonia (Luizão et al. 2007), this toxicity apparently does not affect the occurrence of climax tree species in Guyana (ter Steege and Alexander unpubl. data – in Banki 2010). Additionally, differences in seed sizes influence how tree seedlings respond to soil nutrient conditions (Lawrence 2003). We, therefore, suggest that a more general mechanism explaining habitat specificity should take into account both species characteristics, such as seed size, as well as edaphic properties of these two distinct habitats. So far, the general ecological mechanism driving habitat specificity of tree species to either forest types remains unknown in the upper Rio Negro.
TREE SPECIES DIVERSITY
The high floristic dissimilarity between white-sand and terra-firme forests suggests that these two forest types have a relatively independent evolutionary history. As the evolution of habitat specificity is a long-term process, it ultimately influences the current pattern of tree species diversity. The size of a habitat determines the number of species it can support: large habitats typically host a larger number of species than smaller ones do (Rosenzweig 1995). This prediction holds for various habitats in different geographical regions and is explained by large-scale and long-term evolutionary processes that lead to higher speciation rates and low extinction rates in larger areas (ibid.). Amazonia covers ca. 6.8 million km2 and rainforests extent throughout this area for as long as 55 Million of years (Morley 2000; Jaramillo et al. 2006; Wesselingh et al. 2010). Terra-firme forests cover ca. 80% of the total area of Amazonian forests, whereas white-sand forests cover only 5-10%. The number of species in the regional tree species pool in terra-firme forests is, therefore, expected to be much larger than the regional species pool of trees in white-sand forests (ter Steege et al. 2000; Banki 2010).
The large habitat size of terra-firme forests may also explain the larger number of families, genera, and species associated with these forests. In the presence of two habitats, species may evolve to be habitat specialists or generalists. Assuming that two habitats differ considerably in their size, one would expect to find more habitat-specialist species in larger habitats than in smaller ones (Rosenzweig 1995). If a habitat type occurs in small areas only, species specialized on that habitat may have a disadvantage as they might be more vulnerable to extinction. Hence, we suggest that collectively there might be more species associated with the large terra-firme forests than with the smaller white-sand forests. This hypothesis is supported by Banki (2010) who suggested that terra-firme species experience the most severe establishment limitation in white-sand forests.
How is regional tree diversity related to local diversity? By local diversity we mean the diversity observed at local communities, here analyzed based on one-hectare plots. The local-regional species diversity relationship and the species-area relationship, presented above, are tightly connected. Rosenzweig and Ziv (1999) named this connection the echo pattern. According to the echo pattern, the local-regional species relationship is a direct consequence of the species-area relationship (Figure 6). The connection of these two relationships is strong because species composing the local community are “taken” from the regional community. Therefore, the richer the regional community the richer will be the local community.
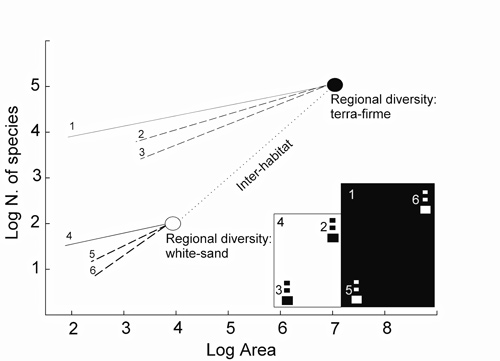
Figure 6 - Theoretical representation of species-area relationships and their connection with the local-regional species relationship (echo pattern: Rosenzweig and Ziv, 1999, Rosenzweig, 1995). The dotted line represents the species-area relationship of distinct habitats, here terra-firme forests (●) and white-sand forests (○). Differences in regional diversity between habitats are mainly explained by evolutionary processes, which are largely influenced by habitat size. Large habitats, such as terra-firme forests (represented by black squares), typically hold a larger number of species than do smaller ones, such as white-sand forests (white squares). The connection between the species-area relationship and the local-regional species relationship is represented by the solid lines. High regional diversity generally causes high local diversity. The dashed lines represent the species-area relationships of ‘islands’, which here represent isolated patches of white-sand or terra-firme forests. Nearby islands (2 and 5) have a less steep slope than islands further away from each other (3 and 6).
How does the echo pattern influence relative species abundance of tree communities in white-sand and terra-firme forests? As white-sand forests have few abundant species in the regional community, few species make up a large part of local community (see Figure 4). Contrary, in the regionally and locally rich terra-firme forest, many species have low abundances; therefore, even the most abundant species in terra-firme forest will have lower abundance than the most abundant species in white-sand forest. In fact, we observed that the four most abundant species accounted for 44% (1,103 trees) of the total number of trees in white-sand forests, whereas this share was only 20% (409 trees) in terra-firme forests.
The low diversity of tree communities of white-sand forests also helps to explain the relatively low floristic dissimilarity of plots within this forest type. Because there are few regionally abundant species in white-sand forests, it is more likely that these few species disperse throughout most patches of this forest type. We found that E. leucantha was the most abundant species in three of the four plots established in white-sand forests, thereby reducing the floristic dissimilarity of tree communities in white-sand forests. The high diversity of tree communities, associated with low relative abundance of dominant tree species, in terra-firme forests resulted in relatively high floristic dissimilarity between individual plots.
FUTURE DIRECTIONS
In our eight one-hectare plots we have collected potentially one new observation for Brazil (Ephedranthus guianensis) (Maas et al. 2010), a first observation for the upper Rio Negro (Pseudoxandra duckei – P. J. M. Maas, pers. comm.), and an undescribed species (Batocarpus sp. – C. C. Berg, pers. comm.). These findings highlight the lacuna we have in our botanical knowledge of the forests of the upper Rio Negro. Deepening our botanical knowledge can be achieved by long-term projects aiming at systematically collecting botanical material (Hopkins 2007). We suggest that establishing long-term monitoring plots designed to capture variation in tree species diversity and composition at multiple spatial scales combined with flora projects planned to collect fertile material of rare species, would make an important contribution to our understanding of tree species diversity and composition in the upper Rio Negro. Long-term projects require long-term collaboration. Our experience shows that collaboration between research institutions and local schools proves to be effective for achieving that aim.
Ackowledgements
We gratefully acknowledge the following colleagues for their support during the field work: Rinaldo and Rose Fernandes, Tone Damito, Luiz Laureano da Silva, Charles Zartman, Laise Diniz, Renata Eiko, students from the ‘Instituto Federal de Educação, Ciência e Tecnologia do Amazonas’ (Ifam - campi SGC and Tunuí), and the Escola Indígena Baniwa Coripaco-Pamaáli (EIBC-Pamaáli). Francimar dos Santos e Joás da Silva provided logistical support in São Gabriel da Cachoeira. The Instituto Socioambiental kindly facilitated the collaboration with EIBC. We acknowledge the invaluable help given by Ana Andrade, Amanda, Alberto Vicentini, Cees C. Berg, Ieda Amaral, Mario Terra, and Paul J. M. Maas in identifying plant material. Michael Hopkins and Carlos Franciscon kindly hosted us at the INPA herbarium. We thank Renata Alves for helping with Figure 1. We thank the Programa BECA (‘Instituto Internacional de Educação do Brasil’ and Gordon and Betty Moore Foundation), National Council of Scientific and Technological Development (CNPq - Brazil), the ‘Alberta Mennega Stichting’, and the ‘Miquel fonds’ for financing the research. We acknowledge OIBI and FUNAI for providing permission to conduct our research at the ‘Terra Indígena do Alto Rio Negro’ (permission No. 0742/07). IBAMA authorized to collect plant material (permission No. 08BR002437/DF). We thank Marinus Werger, Martin Weiss, and three anonymous reviewers for comments on previous versions of this manuscript.
References
Abraao, M.B.; Nelson, B.W.; Baniwa, J.C.; Yu, D.W.; Shepard, G.H. 2008. Ethnobotanical ground-truthing: indigenous knowledge, floristic inventories and satellite imagery in the upper Rio Negro, Brazil. Journal of Biogeography, 35: 2237-2248.
Amaral, I.L.; Matos, F.D.A.; Lima, J. 2000. Composição florística e parâmetros estruturais de um hectare de floresta densa de terra firme no Rio Uatumã, Amazônia, Brasil. Acta Amazonica, 30: 377-392.
Anderson, A.B. 1981. White-sand vegetation of Brazilian Amazonia. Biotropica, 13, 199-210.
ATDN. 2010. Amazon Tree Diversity Network - http://www.bio.uu.nl/~herba/Guyana/ATDN/. Accessed in 12/2010
Banki, O. 2010. Does neutral theory explain community composition in the Guiana shield forests? PhD Thesis. Utrecht University. 148 pp.
Baraloto, C.; Bonal, D.; Goldberg, D.E. 2006. Differential seedling growth response to soil resource availability among nine neotropical tree species. Journal of Tropical Ecology, 22: 487-497.
Bezerra, E.C.; Couto, A.M.R.; Charchar, M.C. 1976. Folha NA. 19 Pico da Neblina; geologia, geomorfologia, pedologia, vegetacao e uso potencial da terra.
Boubli, J.P. 2002. Lowland floristic assessment of Pico da Neblina National Park, Brazil. Plant Ecology, 160: 149-167.
Coomes, D.A.; Grubb, P.J. 1996. Amazonian caatInga and related communities at La Esmeralda, Venezuela: Forest structure, physiognomy and floristics, and control by soil factors. Vegetatio, 122: 167-191.
Costa, F.R.C.; Guillaumet, J.L.; Lima, A.P.; Pereira, O.S. 2009. Gradients within gradients: The mesoscale distribution patterns of palms in a central Amazonian forest. Journal of Vegetation Science, 20: 69-78.
Davis, T.A.W.; Richards, P.W. 1934. The vegetation of moraballi creek, British Guiana: An ecological study of a limited area of tropical rain forest. Part II. Journal of Ecology, 22: 106-155.
Dezzeo, N.; Maquirino, P.; Berry, P ; Aymard, G. 2000. Principales tipos de bosques in el area de San Carlos del Rio Negro. Scientia Guaianae, 11: 15-63.
Dubroeucq, D.; Volkoff, B. 1998. From oxisols to spodosols and histosols: evolution of the soil mantles in the Rio Negro basin (Amazonia). Catena, 32: 245-280.
Dufrene, M.; Legendre, P. 1997. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecological Monographs, 67: 345-366.
Fine, P.V.A.; Mesones, I.; Coley, P.D. 2004. Herbivores promote habitat specialization by trees in Amazonian forests. Science, 305: 663-665.
Fine, P.V.A.; Miller, Z.J.; Mesones, I.;Irazuzta, S.; Appel, H.M.; Stevens, M.H.H.;, Saaksjarvi, I.;Schultz, J.C.; Coley, P.D. 2006. The growth-defense trade-off and habitat specialization by plants in Amazonian forests. Ecology, 87: 150-162.
Fisher, R.A.; Corbet, A.S.; Williams, C.B. 1943. The relation between the number of species and the number of individuals in a random sample of an animal population. Journal of Animal Ecology, 12: 42-58.
Harnik, P.G., Jablonski, D.; Krud, A.Z.; Valentive, J.W. 2010. Genus age, provincial area and the taxonomic structure of marine faunas. Proceedings of the Royal Society B: Biological Sciences, 1 - 9.
Hopkins, M.J.G. 2007. Modelling the known and unknown plant biodiversity of the Amazon Basin. Journal of Biogeography, 34: 1400-1411.
Hubbell, S. P.; He, F.; Condit, R.; Borda de-Água, L.; Kellner, J.; ter Steege, H. 2008. How Many Tree Species Are There in the Amazon and How Many of Them Will Go Extinct? Proceedings of National Academy os Sciences, 105: 11498–11504.
IBGE.1997. Instituto Brasileiro de Geografia e Estatistica. Diagnostico Ambiental da Amazonia Legal. CD-ROM.
Ihaka, R.; Gentleman, R. 1996. R: a language for data analysis and graphics. Journal of computational and graphical statistics, 5: 299-314.
Janzen, D.H. 1974. Tropical blackwater rivers, animals, and mast fruiting by the Dipterocarpaceae. Biotropica, 6: 69-103.
Jaramillo, C.; Rueda, M.J.; Mora, G. 2006. Cenozoic Plant Diversity in the Neotropics. Science, 311: 1893-1896.
Kroonenberg, S.B.; de Roever, E.W.F. 2010. Geological evolution of the Amazonian Craton, p. 9 - 28. In: C. Hoorn & F. Wesselingh (Eds.) Amazonia: landscape and evolution a look into the past. Wiley-Blackwell, Oxford.
Lawrence, D. 2003. The response of tropical tree seedlings to nutrient supply: meta-analysis for understanding a changing tropical landscape. Journal of Tropical Ecology, 19: 239-250.
Lima Filho, D.; Matos, F.D.A., Amaral, I.; Revilla, C.J.; Coelho, L S., Ramos, J.F.; Santos, J.L. 2001. Inventário Florístico de Floresta Ombrófila Densa de Terra Firme, na região do Rio Urucu, Amazonas, Brasil. Acta Amazonica, 31: 565-579.
Luizo, F.J., Luizão, R.C.C.; Proctor, J. 2007. Soil acidity and nutrient deficiency in central Amazonian heath forest soils. Plant Ecology, 192: 209-224.
Maas, J.W.; Rainer, H.; Lobão, A. 2010. Annonaceae in Lista de Espécies da Flora do Brasil. Jardim Botânico do Rio de Janeiro. (http://floradobrasil.jbrj.gov.br/2010/FB110519).
Magurran, A.E. 1988. Ecological Diversity and its measurements. Princeton Univiversity Press, Princeton. 192 pp.
McCune, B.; Grace, J.B.; Urban, D.L. 2002. Analysis of ecological communities. MjM software design Gleneden Beach, Oregon.
McCune, B.; Mefford, M.J. 1999. PC-ORD. Multivariate analysis of ecological data. Version 5.0. MjM Software, Gleneden Beach, Oregon, USA.
Morley, R.J. 2000. Origin and evolution of tropical rain forests. Wiley, New York.
Oliveira, A N.; Amaral, I.; Ramos, M.B.P.; Nobre, A.D.; Couto, L.B.; Sahdo, R.M. 2008. Composição e diversidade florístico-estrutural de um hectare de floresta densa de terra firme na Amazônia Central, Amazonas, Brasil. Acta Amazonica, 38: 627-642.
Oliveira, A.N.; Amaral, I.L. 2004. Florística e fitossociologia de uma floresta de vertente na Amazônia Central, Amazonas, Brasil. Acta Amazonica, 34: 21-34.
Pires, J.M.; Dobzhansky, T.; Black, G.A. 1953. An estimate of the number of species of trees in an Amazonian forest community. Botanical Gazette, 114: 467-477.
Pitman, N.C A.; Terborgh, J.W.; Silman, M.R., Nunez V.P.; Neill, D.A., Ceron, C.E.; Palacios, W.A.; Aulestia, M. 2002. A comparison of tree species diversity in two upper Amazonian forests. Ecology, 83: 3210-3224.
Quesada, C.A. 2008. Soil vegetation interactions across Amazonia. PhD Thesis. School of Geography, University of Leeds. 241 pp.
Rosenzweig, M.L. 1995. Species Diversity in Space and Time. Cambridge University Press, Cambridge. 436 pp.
Rosenzweig, M. L.; Ziv, Y. 1999. The echo pattern of species diversity: pattern and processes. Ecography, 22: 614-628.
Sombroek, W. 2001. Spatial and temporal patterns of Amazon rainfall: Consequences for the planning of agricultural occupation and the protection of primary forests. Ambio, 30: 388-396.
Stropp, J., van der Sleen, P., Quesada, C. A., ter Steege, H. 2011. Seedling traits and herbivory in white-sand and terra-firme forests: an analysis across a resource gradient in the upper Rio Negro, p. 47-69. In: Stropp, J. Towards an understanding of tree diversity in Amazonian forests. PhD Thesis, Utrecht University, Utrecht.
ter Steege, H.; Hammond, D.S. 2001. Character convergence, diversity, and disturbance in tropical rain forest in Guyana. Ecology, 82: 3197-3212.
ter Steege, H.; Sabatier, D.; Castellanos, H.; Van Andel, T.; Duivenvoorden, J.F., De Oliveira, A.A.; Ek, R.; Lilwah, R., Maas, P.; Mori, S.A. 2000. An analysis of the floristic composition and diversity of Amazonian forests including those of the Guiana Shield. Journal of Tropical Ecology, 16: 801-828.
Wesselingh, F.P.; Hoorn, C.; Kroonenberg, S.B.; Antonelli, A.; Lundberg, J.G., Vonhof, H.B.;Hooghiemstra, H. 2010. On the origin of Amazonian landscapes and biodiversity: a synthesis, p. 421 - 432. In C. Hoorn & F. Wesselingh (Eds.) Amazonia, Landscape and Species Evolution. Wiley-Blackwell, Oxford.
Recebido em 10/09/2010
Aceito em 10/03/2011
Appendix: List of all species and morpho-species with total abundance registered in eight one-hectare plots in the upper Rio Negro
Tree species (≥ 10 cm DBH) registered in eight one-hectare plots established in the upper Rio Negro. Species that are significantly associated with one of the two forest types are given in bold (p < 0.05); (*) indicates in the respective column, the forest type the species are associated with (white-sand: WS, terra-firme: TF). The complete dataset of the eight one-hectare plots can be obtained on request to the author.
Family |
Species |
N. trees in WS plots |
N. trees in TF plots |
Anacardiaceae |
Tapirira guianensis Aubl. |
1 |
1 |
Tapirira myriantha Triana & Planch. |
8 |
0 |
|
Thyrsodium guianense Sagot ex Marchand |
0 |
1 |
|
Annonaceae |
Annona sp.1 |
14 |
1 |
Bocageopsis cf. multiflora (Mart.) R.E. Fr. |
0 |
3 |
|
Bocageopsis pleiosperma Maas |
0 |
14* |
|
Diclinanona tessmannii Diels |
0 |
1 |
|
Duguetia sancticaroli Maas |
0 |
4 |
|
Duguetia cf. surinamensis R.E. Fr. |
0 |
1 |
|
Duguetia yeshidan Sandwith |
0 |
1 |
|
Ephedranthus guianensis R.E.Fr. |
0 |
2 |
|
Fusaea longifolia (Aubl.) Saff. |
0 |
8* |
|
Guatteria cf. megalophylla Diels |
1 |
0 |
|
Guatteria guianensis (Aubl.) R.E. Fr. |
0 |
1 |
|
Guatteria insculpta R.E. Fr. |
3 |
0 |
|
Guatteria olivacea R.E. Fr. |
0 |
3 |
|
Guatteria sp.1 |
1 |
0 |
|
Oxandra euneura Diels |
1 |
1 |
|
Pseudoxandra duckei Maas |
0 |
1 |
|
Pseudoxandra sp. |
2 |
0 |
|
Unonopsis duckei R.E. Fr. |
0 |
2 |
|
Xylopia calophylla R.E. Fr. |
0 |
1 |
|
Xylopia cf. emarginata Mart. |
2 |
0 |
|
Xylopia neglecta R.E. Fr. |
1 |
0 |
|
Xylopia spruceana Benth. ex Spruce |
28 |
0 |
|
Xylopia uniflora R.E. Fr. |
31 |
3 |
|
Xylopia sp.1 |
3 |
0 |
|
Xylopia sp.2 |
2 |
0 |
|
Apocynaceae |
Aspidosperma cf. carapanauba Pichon |
1 |
0 |
Aspidosperma nitidum Benth. |
0 |
1 |
|
Aspidosperma obscurinervium Azambuja |
0 |
1 |
|
Aspidosperma sp.1 |
2 |
0 |
|
Aspidosperma sp.2 |
1 |
0 |
|
Aspidosperma sp.3 |
1 |
0 |
|
Couma guianensis Aubl. |
6 |
5 |
|
Couma cf. macrocarpa Barb. Rodr. |
0 |
1 |
|
Couma utilis Müll. Arg. |
1 |
0 |
|
Himatanthus articulatus (Vahl) Woodson |
0 |
1 |
|
Himatanthus obovatus (Müll. Arg.) Woodson |
7 |
1 |
|
Himatanthus sucuuba (Spruce ex Müll. Arg.) Woodson |
2 |
0 |
|
Himatanthus sp.1 |
2 |
0 |
|
Rhigospira quadrangularis (Müll. Arg.) Miers |
20 |
0 |
|
Tabernaemontana macrocalyx Müll. Arg. |
0 |
1 |
|
Tabernaemontana macrophylla Lam. |
1 |
0 |
|
Tabernaemontana sp.1 |
0 |
1 |
|
Indet.1 |
2 |
0 |
|
Araliaceae |
Dendropanax macropodus (Harms) Harms |
2 |
0 |
Dendropanax sp.1 |
1 |
0 |
|
Dendropanax sp.2 |
4 |
0 |
|
Schefflera sp.1 |
0 |
1 |
|
Schefflera sp.2 |
2 |
0 |
|
Arecaceae |
Euterpe precatoria Mart. |
0 |
4 |
Mauritia aculeata Kunth |
2 |
1 |
|
Oenocarpus bacaba Mart. |
0 |
5 |
|
Oenocarpus bataua Mart. |
2 |
1 |
|
Bignoniaceae |
Jacaranda sp.1 |
0 |
3 |
Tabebuia incana A.H. Gentry |
0 |
1 |
|
Boraginaceae |
Cordia exaltata Lam. |
0 |
3 |
Cordia cf. fallax I.M. Johnst. |
0 |
1 |
|
Cordia naidophila I.M. Johnst. |
0 |
1 |
|
Cordia sagotii I.M. Johnst. |
0 |
1 |
|
Burseraceae |
Crepidospermum rhoifolium (Benth.) Triana & Planch |
4 |
2 |
Dacryodes chimatensis Steyerm. & Maguire |
0 |
1 |
|
Dacryodes microcarpa Cuatrec. |
0 |
6 |
|
Indet.1 |
2 |
0 |
|
Indet.2 |
1 |
0 |
|
Protium altsonii Sandwith |
0 |
1 |
|
Protium aff. amazonicum (Cuatrec.) D.C. Daly |
0 |
1 |
|
Protium fimbriatum Swart |
0 |
1 |
|
Protium grandifolium Engl. |
0 |
2 |
|
Protium guianensis (Aubl.) Marchand |
1 |
0 |
|
Protium hebetatum D.C. Daly |
1 |
14 |
|
Protium hebetatum tipo 1 D.C. Daly |
1 |
19 |
|
Protium nitidifolium (Cuatrec.) D.C. Daly |
0 |
2 |
|
Protium paniculatum Engl. |
5 |
3 |
|
Protium robustum (Swart) D.M. Porter |
0 |
3 |
|
Protium spruceanum (Benth.) Engl. |
10 |
2 |
|
Protium sp.1 |
7 |
0 |
|
Protium sp.2 |
2 |
0 |
|
Protium sp.3 |
1 |
1 |
|
Protium sp.4 |
13 |
0 |
|
Protium sp.5 |
0 |
1 |
|
Protium sp.6 |
1 |
2 |
|
Protium sp.7 |
0 |
1 |
|
Protium sp.8 |
2 |
0 |
|
Protium sp.9 |
1 |
0 |
|
Protium sp.10 |
1 |
0 |
|
Protium sp.11 |
18 |
4 |
|
Protium sp.12 |
0 |
1 |
|
Tetragastris panamensis (Engl.) Kuntze |
0 |
2 |
|
Trattinnickia burseraefolia Mart. |
0 |
1 |
|
Trattinnickia glaziovii tipo 1 Swart |
2 |
0 |
|
Trattinnickia glaziovii tipo 2 Swart |
1 |
1 |
|
Trattinnickia sp. |
3 |
1 |
|
Caryocaraceae |
Caryocar glabrum Pers. |
2 |
2 |
Chrysobalanaceae |
Couepia dolichopoda Prance |
0 |
1 |
Couepia edulis (Prance) Prance |
14 |
0 |
|
Couepia elata Ducke |
2 |
2 |
|
Couepia longipendula Pilg. |
1 |
3 |
|
Couepia obovata Ducke |
0 |
1 |
|
Couepia spicata Ducke |
1 |
0 |
|
Couepia sp.1 |
0 |
1 |
|
Hirtella racemosa Lam. |
0 |
1 |
|
Licania aff. kunthiana Hook. f. |
2 |
0 |
|
Licania apetala (E. Mey.) Fritsch |
0 |
1 |
|
Licania bracteata Prance |
0 |
4 |
|
Licania canescens Benoist |
1 |
0 |
|
Licania heteromorpha Benth. |
6 |
3 |
|
Licania kunthiana Hook. f. |
2 |
0 |
|
Licania latifolia Benth. ex Hook. f. |
2 |
0 |
|
Licania laxiflora Fritsch |
9 |
0 |
|
Licania cf. licaniiflora (Sagot) S.F. Blake |
0 |
1 |
|
Licania longistyla (Hook. f.) Fritsch |
4 |
8 |
|
Licania macrophylla Benth. |
2 |
10 |
|
Licania micrantha Miq. |
1 |
5 |
|
Licania octandra (Hoffmanns. ex Roem. & Schult.) Kuntze |
0 |
6 |
|
Licania parviflora Benth. |
15 |
1 |
|
Licania rodriguesii Prance |
0 |
1 |
|
Licania sandwithii Prance |
1 |
0 |
|
Licania sclerophylla (Mart. & Hook. f.) Fritsch |
1 |
3 |
|
Licania sprucei (Hook. f.) Fritsch |
2 |
17 |
|
Licania sp.1 |
5 |
0 |
|
Licania sp.2 |
2 |
6 |
|
Licania sp.3 |
0 |
1 |
|
Parinari sp.1 |
0 |
2 |
|
Clusiaceae |
Calophyllum brasiliense Cambess. |
1 |
0 |
Caraipa densifolia Mart. |
1 |
2 |
|
Caraipa heterocarpa Ducke |
3 |
0 |
|
Caraipa odorata Ducke |
0 |
2 |
|
Caraipa cf. spuria Barb. Rodr. |
4 |
0 |
|
Caraipa sp.1 |
2 |
0 |
|
Caraipa sp.2 |
0 |
13 |
|
Caraipa sp.3 |
1 |
0 |
|
Caraipa sp.4 |
1 |
0 |
|
Caraipa sp.5 |
1 |
0 |
|
Clusia columnaris Engl. |
1 |
0 |
|
Clusia spathulaefolia Engl. |
12 |
0 |
|
Dystovomita brasiliensis D’Arcy |
0 |
6 |
|
Haploclathra paniculata (Mart.) Benth. |
38 |
0 |
|
Symphonia globulifera L. f. |
1 |
7 |
|
Tovomita caloneura A.C. Sm. |
0 |
1 |
|
Tovomita gracilipes Planch. & Triana |
0 |
1 |
|
Tovomita cf. grata Sandwith |
0 |
2 |
|
Tovomita cf. obovata Engl. |
0 |
2 |
|
Tovomita cf. tenuiflora Benth. ex Planch. & Triana |
1 |
0 |
|
Tovomita schomburgkii Planch. & Triana |
2 |
0 |
|
Tovomita sp.1 |
0 |
2 |
|
Tovomita sp.2 |
3 |
0 |
|
Tovomita sp.3 |
0 |
1 |
|
Tovomita umbellata Benth. |
1 |
0 |
|
Vismia cayennensis (Jacq.) Pers. |
0 |
1 |
|
Vismia cf. japurensis Reichardt |
0 |
1 |
|
Indet.1 |
1 |
0 |
|
Combretaceae |
Buchenavia cf. congesta Ducke |
0 |
3 |
Buchenavia oxycarpa (Mart.) Eichler |
0 |
2 |
|
Terminalia sp.1 |
0 |
2 |
|
Indet.1 |
0 |
1 |
|
Cyrillaceae |
Cyrillopsis paraensis Kuhlm. |
1 |
7 |
Dichapetalaceae |
Tapura cf. amazonica Poepp. |
0 |
4 |
Tapura guianensis Aubl. |
1 |
0 |
|
Ebenaceae |
Diospyros aff. bullata A.C. Sm. |
0 |
4 |
Diospyros capreifolia Mart. ex Hiern |
0 |
1 |
|
Diospyros cf. cavalcantei Sothers |
0 |
1 |
|
Diospyros guianensis (Aubl.) Gürke |
1 |
0 |
|
Diospyros sp.1 |
0 |
1 |
|
Elaeocarpaceae |
Sloanea duckei Earle Sm. |
3 |
0 |
Sloanea eichleri K. Schum. |
0 |
1 |
|
Sloanea fendleriana Benth. |
1 |
1 |
|
Sloanea floribunda Spruce ex Benth. |
10 |
0 |
|
Sloanea latifolia (Rich.) K. Schum. |
1 |
0 |
|
Sloanea obtusifolia (Moric.) K. Schum. |
7 |
35 |
|
Sloanea cf. oppositifolia Spruce ex Benth. |
1 |
0 |
|
Sloanea pubescens Benth. |
15 |
14 |
|
Erythroxylum macrophyllum Cav. |
0 |
1 |
|
Erithroxilaceae |
Amanoa aff. guianensis Aubl. |
0 |
1 |
Euphorbiaceae |
Anomalocalyx sp. |
1 |
0 |
Anomalocalyx uleanus (Pax & K. Hoffm.) Ducke |
0 |
1 |
|
Aparisthmium cordatum Baill. |
0 |
3 |
|
Conceveiba guianensis Aubl. |
0 |
5 |
|
Conceveiba martiana Baill. |
1 |
0 |
|
cf. Drypetes variabilis Uittien |
0 |
1 |
|
Hevea benthamiana Müll. Arg. |
6 |
0 |
|
Hevea guianensis Aubl. |
21 |
27 |
|
Micrandra aff. minor Benth. |
23 |
0 |
|
Micrandra spruceana (Baill.) R.E. Schult. |
4 |
71 |
|
Micrandra sprucei (Müll. Arg.) R.E. Schult. |
41 |
1 |
|
Nealchornea sp. |
0 |
1 |
|
Pera bicolor (Klotzsch) Müll. Arg. |
1 |
1 |
|
Pera schomburgkiana (Klotzsch) Müll. Arg. |
0 |
1 |
|
Pera tomentosa (Benth.) Müll. Arg. |
2 |
3 |
|
Pera sp.1 |
0 |
1 |
|
Podocalyx loranthoides Klotzsch |
0 |
5 |
|
Sandwithia guyanensis Lanj. |
0 |
43* |
|
Indet.1 |
1 |
0 |
|
Indet.2 |
1 |
0 |
|
Fabaceae |
Abarema cf. cochleata (Willd.) Barneby & J.W. Grimes |
0 |
2 |
Abarema cf. commutata Barneby & J.W. Grimes |
0 |
5 |
|
Abarema jupunba (Willd.) Britton & Killip |
0 |
3 |
|
Abarema leucophylla (Spruce ex Benth.) Barneby & J.W. Grimes |
8 |
0 |
|
Abarema leucophylla / sp.1 |
4 |
0 |
|
Acosmium cf. nitens (Vogel) Yakovlev |
0 |
1 |
|
Aldina discolor Spruce ex Benth. |
0 |
3 |
|
Aldina cf. macrophylla Spruce ex Benth. |
1 |
0 |
|
Aldina heterophylla Spruce ex Benth. |
137* |
4 |
|
Aldina sp.1 |
6 |
0 |
|
Andira inermis (W. Wright) Kunth ex DC. |
1 |
0 |
|
Andira cf. micans Taub. |
0 |
1 |
|
Andira micrantha Ducke |
5 |
0 |
|
Andira parviflora Ducke |
0 |
1 |
|
Andira unifoliolata Ducke |
2 |
63 |
|
Andira sp.1 |
0 |
3 |
|
Andira sp.2 |
4 |
0 |
|
Andira sp.3 |
9 |
0 |
|
Calliandra tenuiflora Benth. |
0 |
2 |
|
Chamaecrista adiantifolia (Spruce ex Benth.) H.S. Irwin & Barneby |
10 |
3 |
|
Chamaecrista bahiae (H.S. Irwin) H.S. Irwin & Barneby |
1 |
0 |
|
Clathrotropis macrocarpa Ducke |
15 |
138* |
|
Cynometra sp.1 |
0 |
3 |
|
Cynometra sp.2 |
1 |
34 |
|
Dialium guianense (Aubl.) Sandwith |
0 |
4 |
|
Dicymbe pharangiphila R.S. Cowan |
1 |
0 |
|
Dimorphandra coccinea Ducke |
0 |
2 |
|
Diplotropis sp.1 |
0 |
1 |
|
Dipteryx cf. magnifica Ducke |
1 |
3 |
|
Dipteryx rosea Spruce ex Benth. |
0 |
4 |
|
Elizabetha speciosa Ducke |
0 |
1 |
|
Elizabetha sp.1 |
0 |
3 |
|
Eperua leucantha Benth. |
681* |
21 |
|
Eperua purpurea Benth. |
191 |
29 |
|
Hymenaea cf. intermedia Ducke |
0 |
1 |
|
Hymenolobium cf. excelsum Ducke |
0 |
1 |
|
Hymenolobium heterocarpum Ducke |
1 |
0 |
|
Hymenolobium modestum Ducke |
1 |
0 |
|
Inga alba (Sw.) Willd. |
0 |
2 |
|
Inga cordatoalata Ducke |
0 |
1 |
|
Inga lateriflora Miq. |
0 |
1 |
|
Inga leiocalycina Benth. |
0 |
3 |
|
Inga longiflora Spruce ex Benth. |
0 |
1 |
|
Inga marginata Willd. |
0 |
3 |
|
Inga thibaudiana DC. |
0 |
1 |
|
Inga umbratica Poepp. & Endl. |
0 |
1 |
|
Inga sp.1 |
94 |
0 |
|
Inga sp.2 |
5 |
0 |
|
Inga sp.3 |
0 |
2 |
|
Inga sp.4 |
1 |
0 |
|
Macrolobium acaciifolium (Benth.) Benth. |
1 |
0 |
|
Macrolobium angustifolium (Benth.) R.S. Cowan |
26 |
0 |
|
Macrolobium bifolium (Aubl.) Pers. |
23 |
0 |
|
Macrolobium cf. microcalyx Ducke |
4 |
0 |
|
Macrolobium multijugum (DC.) Benth. |
0 |
2 |
|
Macrolobium unijugum Pellegr. |
9 |
3 |
|
Macrolobium sp.1 |
5 |
0 |
|
Macrolobium sp.2 |
12 |
0 |
|
Monopteryx uaucu Spruce ex Benth. |
60 |
74 |
|
Ormosia coarctata Jacks. |
2 |
0 |
|
Ormosia discolor Spruce ex Benth. |
0 |
15 |
|
Ormosia macrophylla Benth. |
2 |
0 |
|
Ormosia cf. paraensis Ducke |
0 |
2 |
|
Ormosia sp.1 |
0 |
1 |
|
Ormosia/Swartzia coarctata/corrugata |
0 |
16 |
|
Parkia igneiflora Ducke |
4 |
0 |
|
Parkia cf. multijuga Benth. |
0 |
2 |
|
Parkia sp.1 |
1 |
0 |
|
Peltogyne campestris Huber ex Ducke |
3 |
0 |
|
Peltogyne cf. excelsa Ducke |
1 |
0 |
|
Peltogyne cf. paniculata Benth. |
0 |
1 |
|
Pseudopiptadenia psilostachya (DC.) G. P. Lewis & M. P. Lima |
1 |
1 |
|
Pterocarpus amazonicus Huber |
2 |
5 |
|
Pterocarpus cf. officinalis Jacq. |
1 |
6 |
|
Pterocarpus rohrii Vahl |
3 |
0 |
|
Pterocarpus santilinoides L’Hér. |
0 |
1 |
|
Pterocarpus sp.1 |
11 |
126 |
|
Sclerolobium chrysophyllum Poepp. |
0 |
1 |
|
Sclerolobium cf. guianense Benth. |
2 |
8 |
|
Sclerolobium sp.1 |
0 |
1 |
|
Slerolobium micropetalum Ducke |
0 |
5 |
|
Stryphnodendron pulcherrimum (Willd.) Hochr. |
0 |
2 |
|
Stryphnodendron sp.1 |
0 |
2 |
|
Swartzia arborescens (Aubl.) Pittier |
0 |
4 |
|
Swartzia cuspidata Spruce ex Benth. |
3 |
16 |
|
Swartzia laevicarpa Amshoff |
1 |
0 |
|
Swartzia polyphylla DC. |
7 |
56 |
|
Swartzia tomentifera (Ducke) Ducke |
3 |
59 |
|
Swartzia sp.1 |
1 |
0 |
|
Swartzia sp.2 |
1 |
0 |
|
Swartzia sp.3 |
2 |
0 |
|
Swartzia sp.4 |
14 |
0 |
|
Swartzia sp.5 |
8 |
0 |
|
Swartzia sp.6 |
2 |
1 |
|
Swartzia sp.7 |
2 |
0 |
|
Swartzia sp.8 |
1 |
0 |
|
Tachigali cf. venusta Dwyer |
0 |
1 |
|
Taralea oppositifolia Aubl. |
48* |
6 |
|
Vatairea guianensis Aubl. |
3 |
0 |
|
Vatairea paraensis Ducke |
0 |
1 |
|
Vatairea sericea (Ducke) Ducke |
1 |
0 |
|
Zygia cf. claviflora (Spruce ex Benth.) Barneby & J.W. Grimes |
0 |
1 |
|
Zygia cf. ramiflora (F. Muell.) Kosterm. |
0 |
1 |
|
Indet.1 |
1 |
0 |
|
Goupiaceae |
Goupia glabra Aubl. |
0 |
8 |
Humiriaceae |
Humiriastrum cuspidatum (Benth.) Cuatrec. |
1 |
1 |
Humiriastrum sp. |
2 |
0 |
|
Hylocarpa heterocarpa (Ducke) Cuatrec. |
16 |
0 |
|
Sacoglottis guianensis Benth. |
2 |
6 |
|
Sacoglottis sp.1 |
1 |
0 |
|
Schistostemon cf. macrophyllum (Benth.) Cuatrec. |
2 |
0 |
|
Schistostemon retusum (Ducke) Cuatrec. |
4 |
0 |
|
Vantanea cf. guianensis Aubl. |
0 |
1 |
|
Vantanea sp.1 |
4 |
0 |
|
Icacinaceae |
Dendrobangia cf. boliviana Rusby |
0 |
1 |
Emmotum aff. nitens (Benth.) Miers |
3 |
0 |
|
Poraqueiba cf. guianensis Aubl. |
1 |
0 |
|
Poraqueiba sericea Tul. |
2 |
1 |
|
Indet. |
Indet.1 |
0 |
1 |
Indet.2 |
1 |
0 |
|
Indet.3 |
1 |
0 |
|
Indet.4 |
0 |
1 |
|
Indet.5 |
0 |
1 |
|
Indet.6 |
1 |
0 |
|
Indet.7 |
1 |
0 |
|
Indet.8 |
1 |
0 |
|
Indet.9 |
0 |
1 |
|
Indet.10 |
0 |
1 |
|
Indet.11 |
1 |
0 |
|
Indet.12 |
1 |
0 |
|
Indet.13 |
0 |
1 |
|
Indet.14 |
1 |
0 |
|
Indet.15 |
0 |
1 |
|
Indet.16 |
1 |
0 |
|
Indet.17 |
0 |
1 |
|
Indet.18 |
1 |
0 |
|
Indet.19 |
0 |
2 |
|
Indet.20 |
0 |
2 |
|
Lauraceae |
Aiouea sp. |
1 |
0 |
Aniba affinis (Meisn.) Mez |
0 |
1 |
|
Aniba williamsii O.C. Schmidt |
0 |
1 |
|
cf. Aiouea or Cinnamomum sp. |
0 |
2 |
|
Licaria cannella (Meisn.) Kosterm. |
0 |
3 |
|
Licaria cf. cannella cannella |
0 |
3 |
|
Licaria guianensis Aubl. |
0 |
3 |
|
Mezilaurus cf. sp. INPA:169.296 |
0 |
1 |
|
Mezilaurus duckei van der Werff |
0 |
1 |
|
Mezilaurus sp.1 |
6 |
0 |
|
Mezilaurus sp.2 |
1 |
0 |
|
Nectandra lanceolata Nees |
0 |
3 |
|
Nectandra cf. lanceolata Nees |
0 |
1 |
|
Nectandra cuspidata Ness |
2 |
0 |
|
Ocotea aciphylla (Nees) Mez |
0 |
2 |
|
Ocotea cernua (Nees) Mez |
0 |
2 |
|
Ocotea cinerea van der Werff |
0 |
9 |
|
Ocotea delicata Vicent. |
0 |
2 |
|
Ocotea cf. douradensis Vatt. |
0 |
2 |
|
Ocotea cf. rhynchophylla (Meisn.) Mez |
3 |
2 |
|
Ocotea cf. tabacifolia (Meisn.) Rohwer |
0 |
2 |
|
Ocotea nitida (Meisn.) Rohwer |
0 |
1 |
|
Ocotea rhodophylla Vicent. |
0 |
1 |
|
Ocotea splendens (Meisn.) Baill |
0 |
1 |
|
Ocotea suaveolens (Meisn.) Benth. & Hook. f. ex Hieron. |
0 |
1 |
|
Ocotea sp.1 |
0 |
17 |
|
Ocotea sp.2 |
0 |
1 |
|
Persea sp. |
1 |
0 |
|
Rhodostemonodaphne crenaticupula Madriñán |
0 |
3 |
|
Rhodostemonodaphne cf. grandis (Mez) Rohwer |
0 |
1 |
|
Rhodostemonodaphne negrensis Madriñán |
1 |
0 |
|
Rhodostemonodaphne sp.1 |
0 |
1 |
|
Sextonia sp.1 |
7 |
0 |
|
Indet.1 |
1 |
0 |
|
Indet.2 |
1 |
0 |
|
Indet.3 |
1 |
0 |
|
Indet.4 |
0 |
1 |
|
Indet.5 |
1 |
0 |
|
Indet.6 |
0 |
1 |
|
Lecythidaceae |
Cariniana micrantha Ducke |
0 |
4 |
Cariniana sp. |
0 |
1 |
|
Couratari gloriosa Sandwith |
0 |
1 |
|
Couratari stellata A.C. Sm. |
0 |
7 |
|
Eschweilera albiflora (DC.) Miers |
0 |
5 |
|
Eschweilera bracteosa (Poepp. ex O. Berg) Miers |
0 |
3 |
|
Eschweilera carinata S.A. Mori |
1 |
7 |
|
Eschweilera decolorans Sandwith |
1 |
1 |
|
Eschweilera gigantea (R. Knuth) J.F. Macbr. |
0 |
6 |
|
Eschweilera cf. macrocarpa Pittier |
0 |
1 |
|
Eschweilera cf. nana (O. Berg) Miers |
0 |
7 |
|
Eschweilera coriacea (DC.) S.A. Mori |
2 |
3 |
|
Eschweilera ovata (Cambess.) Miers |
0 |
2 |
|
Eschweilera pedicellata (Rich.) S.A. Mori |
8 |
55 |
|
Eschweilera pseudodecolorans S.A. Mori |
0 |
1 |
|
Eschweilera revoluta S.A. Mori |
2 |
0 |
|
Eschweilera rionegrense S.A. Mori |
0 |
4 |
|
Eschweilera rodriguesiana S.A. Mori |
0 |
1 |
|
Eschweilera sp.1 |
0 |
1 |
|
Eschweilera subglandulosa (Steud. ex O. Berg) Miers |
0 |
2 |
|
Eschweilera tessmannii R. Knuth |
0 |
11 |
|
Eschweilera wachenheimii (Benoist) Sandwith |
0 |
2 |
|
Gustavia elliptica S.A. Mori |
0 |
2 |
|
Lecythis gracieana S.A. Mori |
0 |
1 |
|
Lecythis parvifructa S.A. Mori |
0 |
6 |
|
Linaceae |
Roucheria punctata (Ducke) Ducke |
7 |
0 |
Malvaceae |
Catostemma sp. |
1 |
0 |
Erisma globosa (Aubl.) A.Robyns |
8 |
2 |
|
Pachira cf. aquatica Aubl. |
2 |
0 |
|
Pachira aff. insignis (Sw.) Sw. ex Savigny |
1 |
0 |
|
Pachira coriacea (Mart.) W.S. Alverson |
1 |
0 |
|
Pachira sp.1 |
2 |
0 |
|
Quararibea ochrocalyx (K. Schum.) Vischer |
0 |
1 |
|
Scleronema micranthum (Ducke) Ducke |
43 |
24 |
|
Theobroma subincanum Mart. |
0 |
8 |
|
Theobroma sp. |
0 |
1 |
|
Melastomataceae |
Miconia lepidota Schrank & Mart. ex DC. |
0 |
3 |
Miconia punctata (Desr.) D. Don ex DC. |
0 |
2 |
|
Miconia pyrifolia Naudin |
0 |
4 |
|
Mouriri ficoides Morley |
0 |
5 |
|
Meliaceae |
Carapa cf. guianensis Aubl. |
0 |
2 |
Guarea convergens T.D. Penn. |
0 |
4 |
|
Guarea trunciflora C. DC. |
0 |
2 |
|
Trichilia sp.1 |
1 |
0 |
|
Trichilia sp.2 |
0 |
2 |
|
Monimiaceae |
Siparuna decipiens (Tul.) A. DC. |
0 |
6 |
Siparuna guianensis Aubl. |
0 |
1 |
|
Siparuna poeppigii (Tul.) A. DC. |
1 |
1 |
|
Moraceae |
Batocarpus sp. |
0 |
1 |
Brosimum acutifolium Huber |
2 |
1 |
|
Brosimum guianense Huber ex Ducke |
4 |
8 |
|
Brosimum lactescens (S.Moore) C.C. Berg |
1 |
13* |
|
Brosimum melanopotamicum C.C. Berg |
1 |
1 |
|
Brosimum ovatifolium Ducke |
2 |
0 |
|
Brosimum rubescens Taub. |
6 |
16 |
|
Brosimum sp.1 |
1 |
0 |
|
Brosimum sp.3 |
0 |
1 |
|
Brosimum utile (Kunth) Oken ex J. Presl |
1 |
4 |
|
Clarisia sp.1 |
0 |
1 |
|
Ficus americana Aubl. |
1 |
0 |
|
Ficus paraensis (Miq.) Miq. |
0 |
1 |
|
Helianthostylis sprucei Baill. |
0 |
6 |
|
Helianthostylis steyermarkii C.C. Berg |
0 |
6 |
|
Helicostylis scabra (J.F. Macbr.) C.C. Berg |
1 |
4 |
|
Helicostylis sp.1 |
0 |
1 |
|
Helicostylis tomentosa (Poepp. & Endl.) Rusby |
0 |
8 |
|
Maquira sp.1 |
0 |
1 |
|
Naucleopsis oblongifolia (Kuhlm.) Carauta |
0 |
4 |
|
Pseudolmedia macrophylla Trécul |
0 |
1 |
|
Trymatococcus amazonicus Poepp. & Endl. |
0 |
5 |
|
Myristicaceae |
Iryanthera coriacea Ducke |
21 |
2 |
Iryanthera crassifolia A.C. Sm. |
0 |
2 |
|
Iryanthera elliptica Ducke |
0 |
20* |
|
Iryanthera laevis Markgr. |
14 |
26 |
|
Iryanthera lorentensis/lancifolia |
5 |
6 |
|
Iryanthera olacoides (A.C. Sm.) A.C. Sm. |
0 |
2 |
|
Iryanthera paraensis Huber |
0 |
5 |
|
Iryanthera sagotiana (Benth.) Warb. |
0 |
1 |
|
Iryanthera ulei Warb. |
5 |
49* |
|
Iryanthera/Osteophloeum sagotiana/platyspermum |
11 |
12 |
|
Iryanthera sp.1 |
0 |
2 |
|
Osteophloeum platyspermum (Spruce ex A. DC.) Warb. |
0 |
1 |
|
Otoba parvifolia (Markgr.) A.H. Gentry |
0 |
2 |
|
Otoba sp.1 |
1 |
0 |
|
Virola calophylla/pavonis |
0 |
50 |
|
Virola macrocarpa A.C. Sm. |
0 |
2 |
|
Virola michelii Heckel |
2 |
7 |
|
Virola minutiflora Ducke |
0 |
1 |
|
Virola mollissima (Poepp. ex A. DC.) Warb. |
0 |
2 |
|
Virola cf. theiodora (Spruce ex Benth.) Warb. |
0 |
3 |
|
Virola surinamensis (Rol. ex Rottb.) Warb. |
0 |
9 |
|
Virola sp.1 |
0 |
6 |
|
Virola sp.2 |
2 |
7 |
|
Virola sp.3 |
2 |
3 |
|
Myrsinaceae |
Cybianthus fulvopulverulentus (Mez) G. Agostini |
2 |
0 |
Myrtaceae |
Calycolpus sp. |
0 |
1 |
Calyptranthes cuspidata DC. |
0 |
1 |
|
Calyptranthes densiflora Poepp. ex O. Berg |
0 |
1 |
|
Campomanesia sp.1 |
0 |
2 |
|
Eugenia diplocampta Diels |
0 |
1 |
|
Eugenia patrisii Vahl |
0 |
1 |
|
Myrcia magnoliifolia (O. Berg) Kiaersk. |
0 |
1 |
|
Myrcia sp.1 |
1 |
0 |
|
Myrcia sp.2 |
1 |
0 |
|
Nyctaginaceae |
Guapira sancarlosiana Steyerm. |
1 |
0 |
Neea cf. obovata 2 Spruce ex Heimerl |
2 |
0 |
|
Neea madeirana Standl. |
2 |
0 |
|
Neea oppositifolia Ruiz & Pav. |
5 |
0 |
|
Neea ovalifolia Spruce ex J.A. Schmidt |
0 |
5 |
|
Neea robusta Steyerm. |
2 |
2 |
|
Neea sp.1 |
0 |
1 |
|
Neea sp.5 |
1 |
0 |
|
Pisonia tomentosa Casar. |
0 |
1 |
|
Indet.1 |
0 |
1 |
|
Ochnaceae |
Ouratea discophora Ducke |
1 |
0 |
Olacaceae |
Cathedra sp.1 |
0 |
1 |
Dulacia guianensis (Engl.) Kuntze |
0 |
1 |
|
Heisteria densifrons Engl. |
0 |
4 |
|
Heisteria duckei Sleumer |
0 |
1 |
|
Heisteria krukovii A.C. Sm. |
0 |
1 |
|
Minquartia guianensis Aubl. |
1 |
12 |
|
Quiinaceae |
Lacunaria crenata (Tul.) A. C. Smith |
0 |
2 |
Lacunaria cf. jenmanii (Oliv.) Ducke |
0 |
2 |
|
Lacunaria sp.1 |
0 |
1 |
|
Quiina sp. |
0 |
1 |
|
Rhabdodendraceae |
Rhabdodendron amazonicum (Spruce ex Benth.) Huber |
11 |
0 |
Rubiaceae |
Botryarrhena pendula Ducke |
0 |
5 |
Calycophyllum acreanum Ducke |
1 |
0 |
|
Capirona sp.1 |
0 |
1 |
|
Chimarrhis duckeana Delprete |
0 |
14* |
|
Chimarrhis sp.1 |
0 |
1 |
|
Duroia cf. macrophylla Huber |
0 |
4 |
|
Duroia saccifera (Mart. ex Roem. & Schult.) Hook. f. ex Schumann |
0 |
2 |
|
Duroia velutina Hook. f. ex K. Schum. |
0 |
1 |
|
Ferdinandusa uaupensis Spruce ex K. Schum. |
4 |
1 |
|
Isertia hypoleuca Benth. |
0 |
1 |
|
Libertia sp.1 |
0 |
1 |
|
Pagamea acrensis Steyerm. |
1 |
0 |
|
Pagamea coriacea Spruce ex Benth. |
4 |
0 |
|
Pagamea cf. macrophylla Spruce ex Benth. |
5 |
1 |
|
Pagamea plicata Spruce ex Benth. |
20* |
0 |
|
Palicourea amapaensis Steyerm. |
0 |
1 |
|
Psychotria sp.1 |
0 |
1 |
|
Remijia hispida Spruce ex K. Schum. |
1 |
0 |
|
Remijia aff. ulei K. Krause |
0 |
2 |
|
Remijia sp.1 |
0 |
1 |
|
Warszewiczia schwackei K. Schum. |
0 |
1 |
|
Rutaceae |
Hortia longifolia Benth. ex Engl. |
3 |
0 |
Sapindaceae |
Cupania emarginata Cambess. |
2 |
0 |
Cupania hispida Radlk. |
0 |
1 |
|
Cupania scrobiculata Rich. |
0 |
1 |
|
Matayba elegans Radlk. |
0 |
1 |
|
Talisia pachycarpa Radlk. |
0 |
1 |
|
Talisia sp. |
0 |
1 |
|
Sapotaceae |
Chromolucuma rubriflora Ducke |
0 |
2 |
Chrysophyllum amazonicum T.D. Penn. |
0 |
4 |
|
Chrysophyllum cuneifolium (Rudge) A. DC. |
0 |
1 |
|
Chrysophyllum pomiferum (Eyma) T.D. Penn. |
0 |
10 |
|
Chrysophyllum prieurii A. DC. |
1 |
4 |
|
Chrysophyllum sanguinolentum (Pierre) Baehni |
19 |
9 |
|
Chrysophyllum sp.1 |
0 |
1 |
|
Chrysophyllum sp.2 |
0 |
1 |
|
Ecclinusa guianensis Eyma |
0 |
1 |
|
Ecclinusa sp.1 |
1 |
2 |
|
Ecclinusa sp.2 |
0 |
1 |
|
Ecclinusa sp.3 |
0 |
1 |
|
Manilkara cf. amazonica (Huber) A. Chev. |
15 |
3 |
|
Manilkara bidentata (A. DC.) A. Chev. |
13 |
0 |
|
Manilkara bidentata/cuspidata |
8 |
0 |
|
Manilkara sp.1 |
5 |
0 |
|
Manilkara sp.2 |
1 |
0 |
|
Micropholis cf. acutangula (Ducke) Eyma |
0 |
1 |
|
Micropholis cylindrocarpa (Poepp. & Endl.) Pierre |
0 |
1 |
|
Micropholis cf. grandiflora Aubrév. |
0 |
1 |
|
Micropholis guyanensis (A. DC.) Pierre |
3 |
32 |
|
Micropholis splendens Gilly ex Aubréville |
0 |
4 |
|
Micropholis trunciflora Ducke |
1 |
0 |
|
Micropholis venulosa (Mart. & Eichler) Pierre |
1 |
5 |
|
Micropholis sp.1 |
0 |
2 |
|
Pouteria cf. ambelaniifolia (Sandwith) T.D. Penn. |
0 |
1 |
|
Pouteria cf. cladantha Sandwith |
5 |
2 |
|
Pouteria cuspidata (A. DC.) Baehni |
6 |
5 |
|
Pouteria elegans (A. DC.) Baehni |
8 |
1 |
|
Pouteria eugenifolia (Pierre) Baehni |
0 |
1 |
|
Pouteria filipes Eyma |
0 |
1 |
|
Pouteria fimbriata Baehni |
1 |
1 |
|
Pouteria hispida Eyma |
0 |
4 |
|
Pouteria cf. latianthera T.D. Penn. |
0 |
1 |
|
Pouteria opposita (Ducke) T.D. Penn. |
0 |
1 |
|
Pouteria sp.1 |
0 |
2 |
|
Pouteria sp.2 |
0 |
1 |
|
Pouteria sp.3 |
1 |
0 |
|
Pouteria sp.4 |
1 |
0 |
|
Pouteria sp.5 |
6 |
0 |
|
Pouteria sp.6 |
25 |
0 |
|
Pouteria sp.7 |
0 |
1 |
|
Pouteria sp.8 |
0 |
1 |
|
Pouteria sp.9 |
1 |
0 |
|
Pouteria sp.10 |
1 |
0 |
|
Pouteria sp.11 |
0 |
1 |
|
Pradosia beardii (Monach.) T.D. Penn. |
1 |
0 |
|
Pradosia cf. mutisii Cronquist |
47* |
0 |
|
Pradosia schomburgkiana (A. DC.) Cronquist |
2 |
3 |
|
Pradosia sp.1 |
2 |
0 |
|
Sterculiaceae |
Sterculia sp.1 |
0 |
4 |
Tiliaceae |
Apeiba echinata Gaertn. |
0 |
5 |
Apeiba sp.1 |
0 |
1 |
|
Ulmaceae |
Ampelocera ruizii Klotzsch |
3 |
0 |
Ampelocera sp.1 |
0 |
2 |
|
Ampelocera sp.2 |
1 |
1 |
|
Urticaceae |
Cecropia cf. latiloba Miq. |
0 |
1 |
Cecropia cf. sciadophylla Mart. |
0 |
2 |
|
Cecropia purpurascens C.C. Berg |
0 |
6 |
|
Pourouma bicolor Mart. |
0 |
2 |
|
Pourouma minor Benoist |
0 |
3 |
|
Pourouma sp.1 |
0 |
1 |
|
Pourouma tomentosa Mart. ex Miq. |
13 |
2 |
|
Verbenaceae |
Vitex calothyrsa Sandwith |
29 |
0 |
Vitex orinocensis Kunth |
0 |
3 |
|
Violaceae |
Rinorea falcata (Mart. ex Eichler) Kuntze |
0 |
1 |
Vochysiaceae |
Erisma bicolor Ducke |
1 |
0 |
Erisma calcaratum (Link) Warm. |
0 |
34 |
|
Erisma japura Spruce ex Warm. |
0 |
3 |
|
Erisma sp.1 |
0 |
1 |
|
Ruizterania cassiquiarensis (Spruce ex Warm.) Marc.-Berti |
9 |
0 |
|
Ruizterania sp.1 |
6 |
4 |
|
Vochysia bicolor Ducke |
1 |
0 |
|
Vochysia spatiphylla Stafleu |
0 |
1 |
|
Vochysia vismiifolia Spruce ex Warm. |
0 |
4 |
|
Vochysia sp.1 |
0 |
3 |
|
Vochysia sp.2 |
0 |
1 |
|
|
Total |
2524 |
2179 |